Zhi Zhang’s team at USTC provides a detailed analysis of the core regulatory circuit in Science
Release time:2026-05-09 14:49:37
Human pain sensitivity exhibits daily fluctuations: patients with neuropathic pain or rheumatoid arthritis often experience significantly heightened pain in the evening. However, the underlying neural mechanisms remain unclear. Although the central nervous system shows rhythmic activity, direct evidence linking neuronal circadian rhythms to pain sensitivity is limited. The daytime activity patterns of pain-processing brain regions under pathological pain remain undefined, and how the central master clock coordinates these pain-related regions is still unknown.
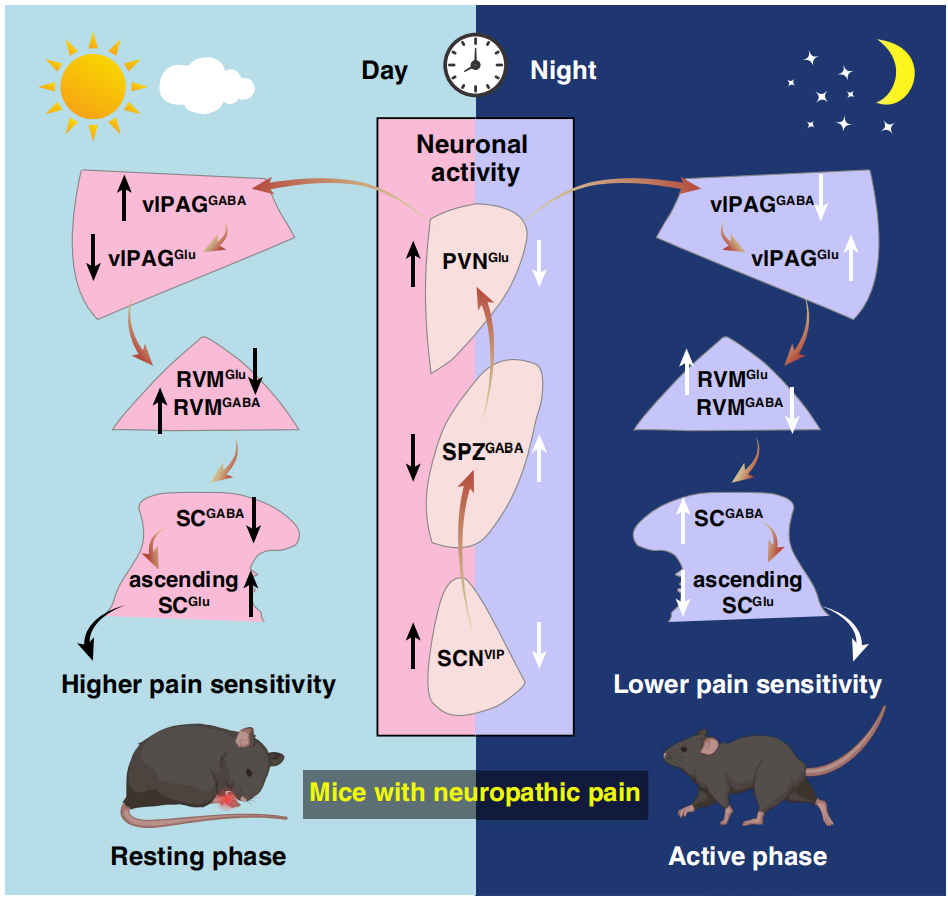
On March 19, 2026, Zhi Zhang, Yan Jin, and Junchao Qian’s team at the University of Science and Technology of China published a paper in Science titled “Hypothalamic clock governs circadian pain”, revealing for the first time the neural circuit mechanisms by which the hypothalamic master clock regulates the circadian rhythm of chronic pain. They found that neuropathic pain mice exhibited higher pain sensitivity during the day (rest phase, ZT5) and lower sensitivity at night (active phase, ZT14). This rhythm is driven by a polysynaptic SCNVIP → SPZGABA → PVNGlu → vlPAGGABA circuit and mediated through the descending analgesic pathway from vlPAG → RVM → spinal cord. SCN VIP neurons act as the central regulatory node, with SPZ serving as a key inhibitory relay. This discovery provides a novel mechanistic framework and potential targets for time-based therapy of chronic pain.
Circadian Rhythm of Nociceptive Sensitivity in Pain Mouse Models
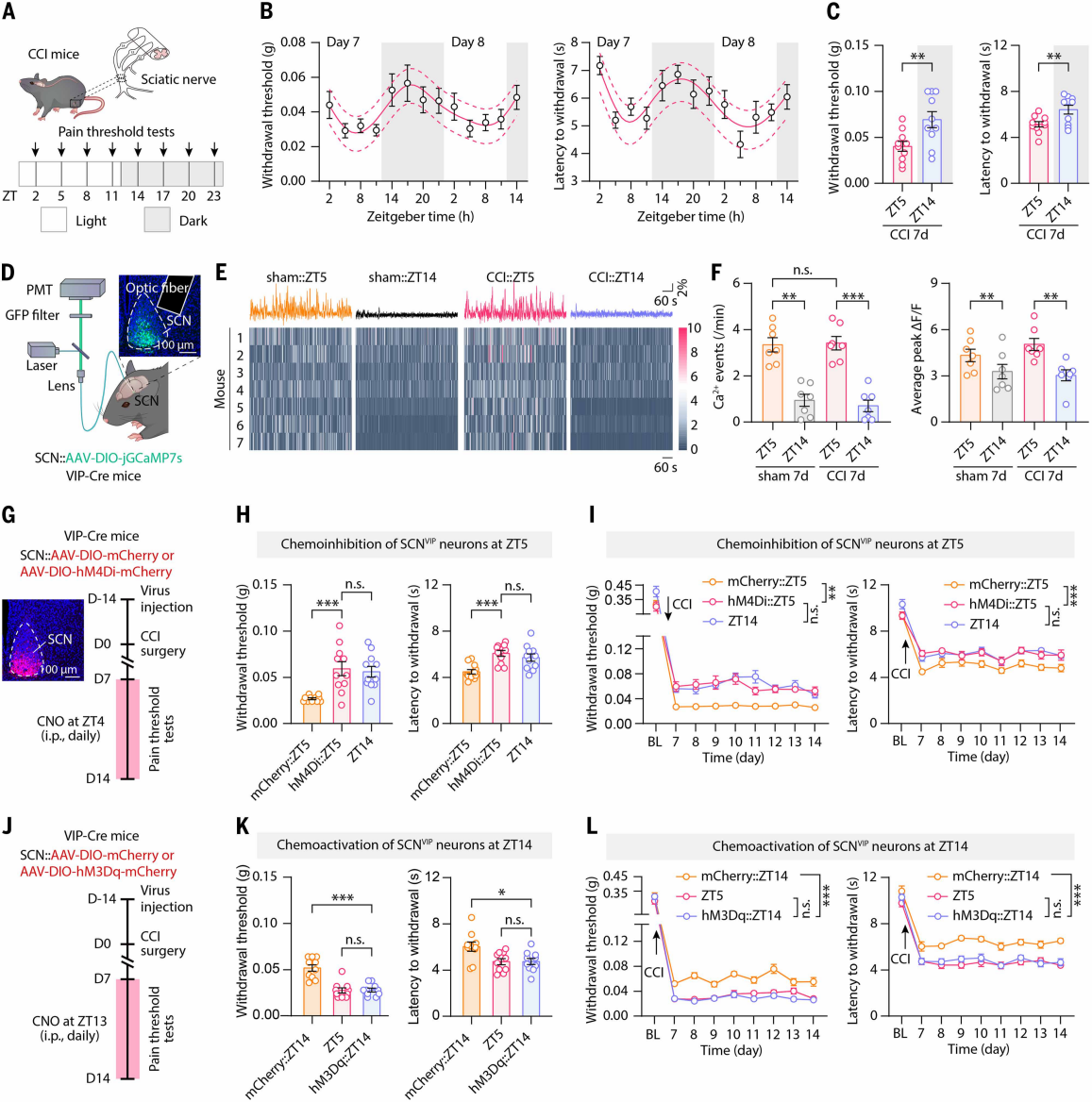
Mice are primarily active at night and rest during the day. To investigate circadian differences in pain sensitivity, the authors established a neuropathic pain model using chronic constriction injury (CCI) of the sciatic nerve. Under a 12-hour light-dark cycle, mechanical and thermal nociceptive thresholds were measured at eight time points (ZT2, ZT5, etc.) (Figure 1A). Results showed that both thresholds fluctuated in a circadian manner, peaking at ZT17 and reaching a trough at ZT5 (Figure 1B). On post-CCI day 7, mice exhibited significantly higher pain thresholds at the first testing point in the dark phase (ZT14) than at ZT5 (Figure 1C), so subsequent experiments used ZT14 for measurements. The circadian rhythm of pain thresholds persisted for at least 14 days post-CCI. A similar trend was observed in the inflammatory pain model induced by complete Freund’s adjuvant (CFA) injected into the hindpaw, indicating that mice with neuropathic or inflammatory pain are more sensitive to pain during the daytime (rest phase) and less sensitive at night (active phase). Placing CCI mice under constant darkness to eliminate light cues still produced circadian variations in pain sensitivity, although the ratio of pain thresholds at CT14 to CT5 decreased, suggesting that this rhythm is driven by both exogenous light and endogenous circadian mechanisms.
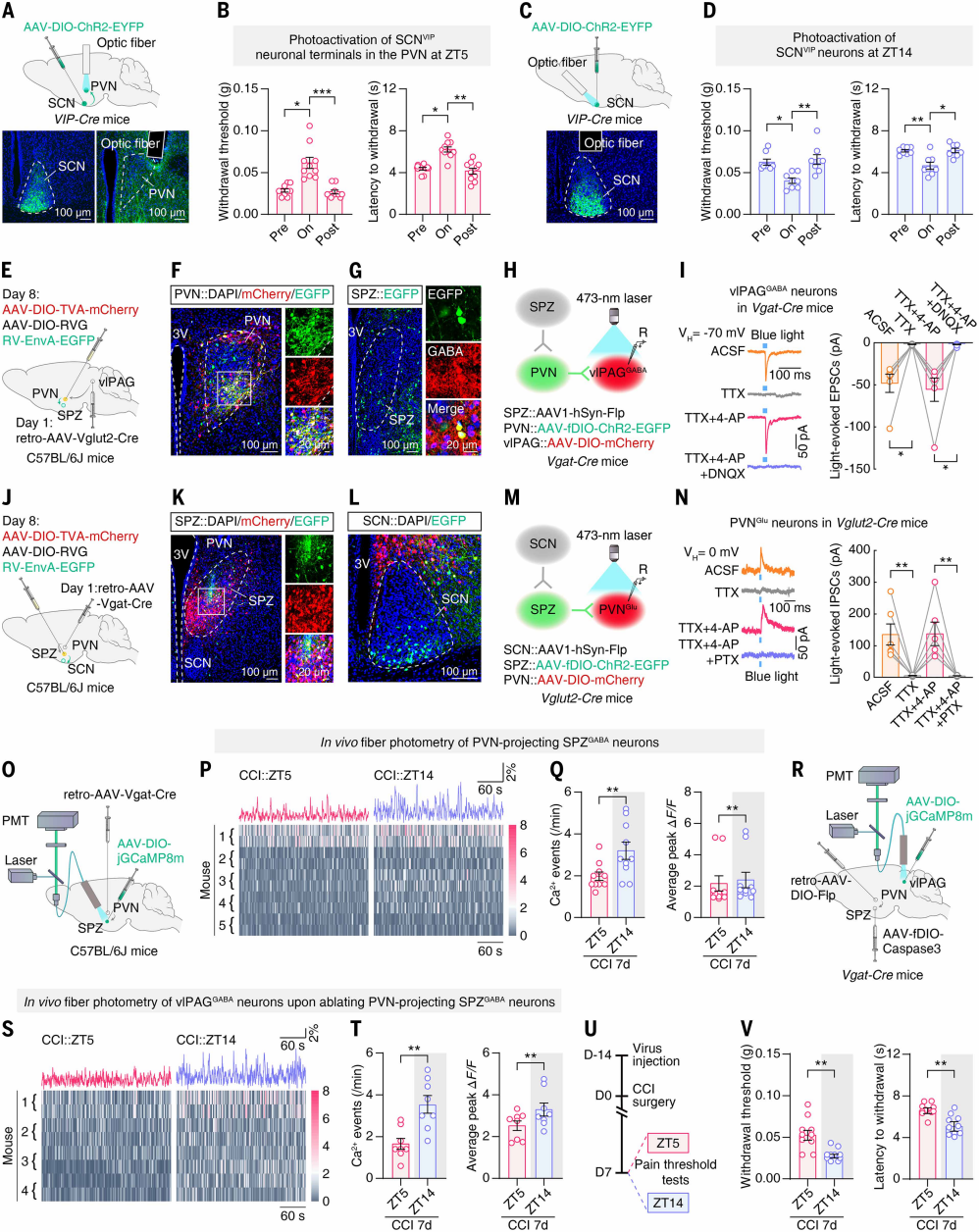
Circadian Activity of SCNVIP Neurons Mediates Pain Rhythms
Given the established role of the hypothalamic suprachiasmatic nucleus (SCN) as the central circadian clock, the authors examined whether SCN activity correlates with pain behavior rhythms. Following previous studies, CCI day 7 mice received SCN injections of AAV-jGCaMP7s. In these mice, SCN neuronal Ca²⁺ events were more frequent and peaked higher during the day (ZT2, ZT5) and reduced at night (ZT14). Both Ca²⁺ activity and the number of Fos-TRAP-labeled neurons at ZT5 were higher than at ZT14, indicating that SCN neuronal activity fluctuates in parallel with pain sensitivity.
SCN neurons expressing vasoactive intestinal peptide (SCNVIP) and arginine vasopressin (SCNAVP) regulate circadian behavior. After AAV-DIO-jGCaMP7s injections, both SCNVIP and SCNAVP neurons exhibited higher Ca²⁺ event frequency and peak values at ZT5 than at ZT14 (Figures 1D-F). To test their functional role, VIP-Cre and AVP-Cre mice received SCN injections to inhibit or activate these neurons (Figures 1G, 1J). Chemogenetic inhibition of SCNVIP neurons at ZT4 elevated the ZT5 pain threshold to near ZT14 levels (Figure 1H), while activation at ZT14 lowered pain thresholds to ZT5 levels (Figure 1K). Continuous inhibition or activation produced similar effects (Figures 1I, 1L). Manipulation of SCNAVP neurons did not affect circadian pain fluctuations. Collectively, rhythmic activity of SCNVIP neurons is necessary for the circadian pain sensitivity of CCI mice.
Figure 1. Circadian pain sensitivity in CCI mice requires rhythmic SCN activity
vlPAGGABA Neuron Activity Synchronizes with Circadian Pain
Because the SCN is not a classical pain-processing region, the authors next screened pain-related brain areas for rhythmic activity. Fos-TRAP labeling revealed that, compared with sham controls, CCI mice exhibited more tdTomato⁺TRAP neurons in multiple brain regions, including mPFC, ACC, CeA, lPAG, and vlPAG. Within CeA, lPAG, vlPAG, and DRN, the number of tdTomato⁺TRAP-labeled neurons was significantly higher at ZT5 than ZT14, indicating higher neuronal responsiveness during the day.
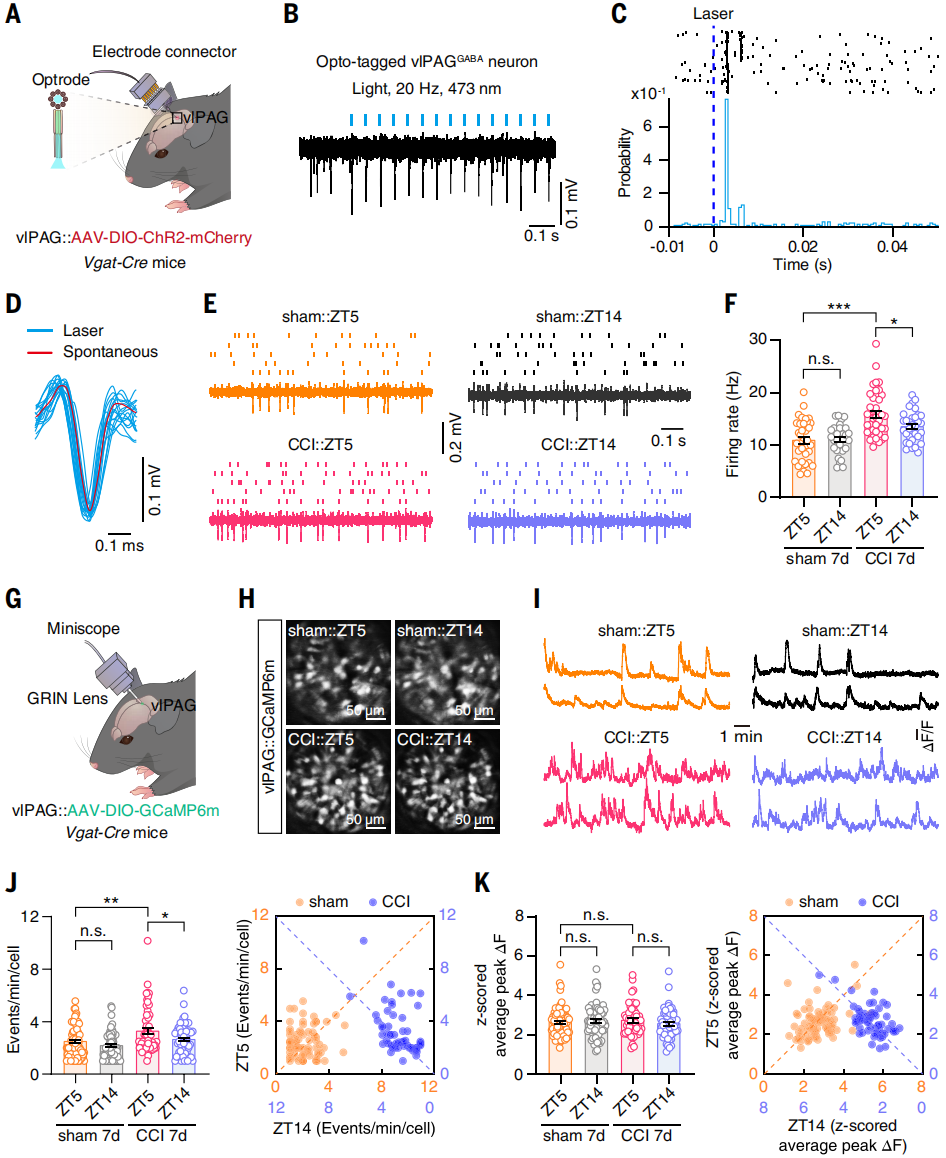
The vlPAG showed the lowest ZT14/ZT5 ratio of tdTomato⁺ neurons and is a known component of the descending analgesic pathway. The authors therefore examined whether vlPAG contributes to circadian changes in nociceptive sensitivity in neuropathic pain mice. Since vlPAG GABAergic neurons (vlPAGGABA) are known to facilitate nociceptive responses, their activity rhythms were evaluated using in vivo multi-electrode recording and miniature microscope Ca²⁺ imaging in freely moving Vgat-Cre mice. Analysis of spike waveforms evoked by laser stimulation (473 nm, 20 Hz) revealed that firing rates of vlPAGGABA neurons were higher at ZT5 than ZT14 in CCI day 7 mice (Figures 2A-F).
Furthermore, Vgat-Cre mice injected with AAV-DIO-GCaMP6m into vlPAG and imaged with a miniature microscope showed elevated Ca²⁺ transient frequency in CCI mice compared with sham controls, with higher activity at ZT5 than ZT14 (Figures 2G-K). These results indicate that the rhythmic activity of vlPAGGABA neurons is synchronized with the circadian fluctuation of pain sensitivity.
Figure 2. Circadian activity of vlPAGGABA neurons in CCI mice
Rhythmic Activity of the PVNGlu → vlPAGGABA Circuit Mediates Circadian Pain
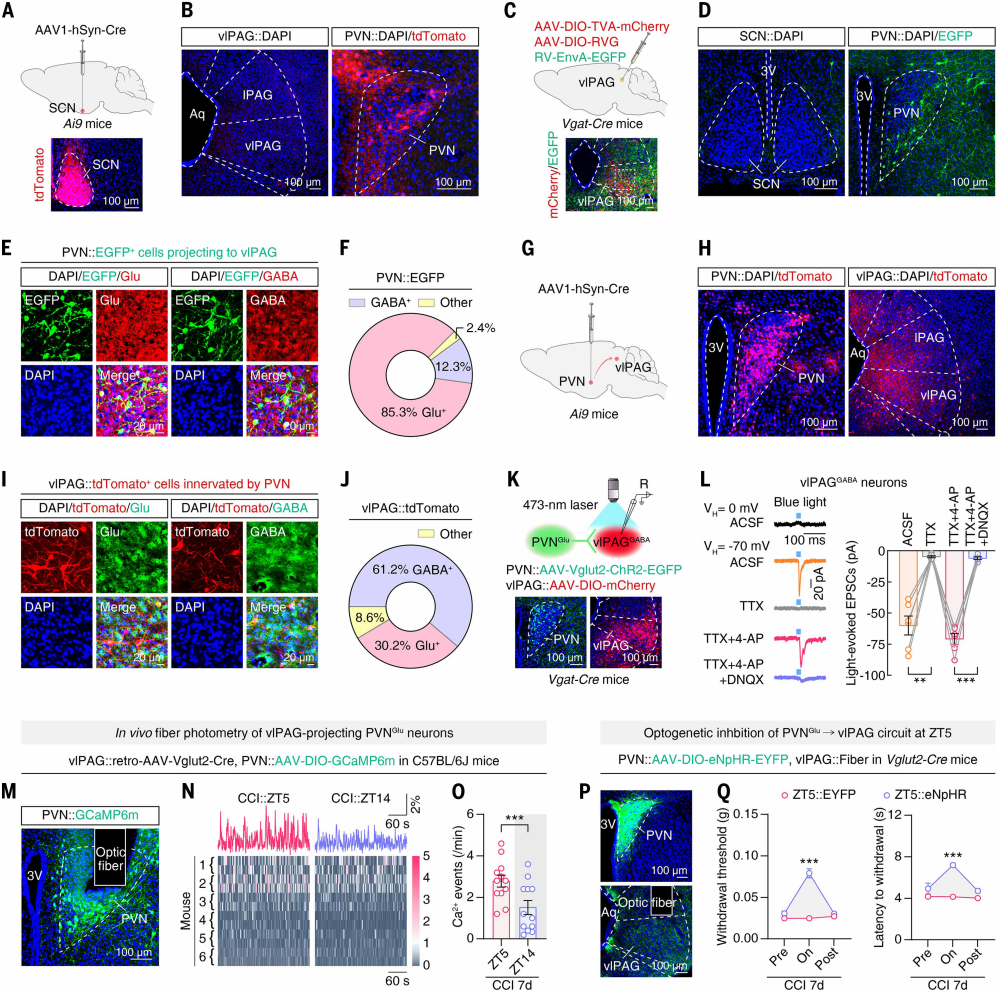
The authors investigated whether the rhythmic activity of vlPAGGABA neurons in CCI mice is directly driven by SCN input. Anterograde trans-synaptic tracing with AAV1-hSyn-Cre was injected into the SCN of Ai9 mice (Figure 3A). tdTomato⁺ neurons were observed in SPZ, PVN, and other brain regions, but no signal was detected in vlPAG or classical pain-related areas (Figure 3B), indicating that the SCN does not directly project to vlPAG.
To identify SCN downstream regions projecting directly to vlPAG, retrograde tracing was performed by injecting helper virus and RV virus into the vlPAG of Vgat-Cre mice (Figure 3C). Among SCN’s main downstream targets, PVN showed the most EGFP⁺ signal (Figure 3D), 85.3% of which co-localized with glutamate immunostaining (Figures 3E-F). Anterograde tracing from PVN in Ai9 mice revealed that 61.2% of tdTomato⁺ neurons in vlPAG co-stained with GABA antibodies (Figures 3G-J).
To confirm functional synaptic connectivity of the PVNGlu → vlPAGGABA circuit, PVN was injected with AAV-Vglut2-ChR2-EGFP and vlPAG of Vgat-Cre mice with AAV-DIO-mCherry (Figure 3K). Whole-cell patch-clamp recordings in brain slices showed that photostimulation of PVNGlu terminals evoked EPSCs in vlPAGGABA neurons, which were abolished by TTX, recovered by TTX + 4-AP, and almost completely blocked by DNQX (AMPA receptor antagonist) (Figure 3L), confirming monosynaptic glutamatergic connectivity.
TRAP2;Ai14 labeling on CCI day 7 mice showed more PVN tdTomato⁺ neurons at ZT5 than ZT14. Following injection of retrograde AAV-Vglut2-Cre into vlPAG and AAV-DIO-GCaMP6m into PVN, fiber photometry revealed higher Ca²⁺ event frequency of PVNGlu neurons at ZT5 compared with ZT14 in CCI mice; sham controls showed no difference (Figures 3M-O), indicating circadian rhythmicity. Optogenetic manipulation showed that photoinhibition of PVNGlu terminals in vlPAG at ZT5 (via AAV-DIO-eNpHR-EYFP injection into PVN) increased pain thresholds, whereas activation at ZT14 decreased pain thresholds (Figures 3P-Q). These results demonstrate that rhythmic activity of the PVNGlu → vlPAGGABA circuit drives circadian pain sensitivity in CCI mice.
Figure 3. Rhythmic activity of the PVNGlu → vlPAGGABA circuit mediates circadian pain
SPZ as an Inhibitory Relay Between SCN and PVN
The SCN is mainly composed of inhibitory GABAergic neurons but can also exert excitatory effects. Previous studies show that SCN electrical stimulation induces monosynaptic inhibitory and excitatory postsynaptic potentials in PVN neurons. To test whether higher daytime pain sensitivity in CCI mice results from direct excitatory SCN input to PVN, the authors injected AAV-DIO-ChR2-EYFP into SCN and implanted optical fibers in PVN of CCI day 7 VIP-Cre mice for optogenetic and behavioral testing (Figure 4A). Activation of SCNVIP terminals in PVN at ZT5 increased pain thresholds and reduced sensitivity (Figure 4B); direct SCNVIP activation at ZT14 decreased pain thresholds (Figures 4C-D). Similar results were obtained in Vgat-Cre mice when SCN GABAergic neurons were activated. These findings suggest that SCN not only sends direct inhibitory signals to PVN but may also exert disinhibitory effects via intermediate inhibitory brain regions, synchronizing neuronal activity.
To examine how SCN downstream inhibitory nuclei regulate the PVNGlu → vlPAGGABA circuit, a triple retrograde tracing approach was used: after retrograde AAV-Vglut2-Cre injection into vlPAG, PVN was injected with helper virus and RV-ΔG-EGFP (Figures 4E-F). EGFP⁺ cells were detected in SPZ (the main downstream target of SCN, composed of GABAergic neurons), indicating that SPZGABA neurons control PVNGlu neurons projecting to vlPAG. Viral injections into SPZ, PVN, and vlPAG in Vgat-Cre mice followed by brain slice recordings revealed that blue-light stimulation evoked DNQX-sensitive EPSCs in mCherry⁺ vlPAGGABA neurons (Figures 4H-I), confirming monosynaptic excitatory projections from SPZ-controlled PVNGlu neurons. Retrograde monosynaptic tracing also showed abundant EGFP⁺ SCN cells after PVN injection of retrograde AAV-Vgat-Cre and SPZ injection of the corresponding virus (Figures 4J-L), indicating SCN innervation of SPZGABA neurons projecting to PVN. In Vglut2-Cre mice, photostimulation of ChR2⁺ PVN terminals evoked IPSCs in mCherry⁺ PVNGlu neurons, which were abolished by PTX (Figures 4M-N), confirming monosynaptic inhibitory projections from SPZGABA neurons to PVNGlu neurons. Collectively, these results delineate the anatomical connectivity of SPZGABA → PVNGlu → vlPAGGABA and SCN → SPZGABA → PVN circuits, establishing SPZ as an inhibitory relay between SCN and PVNGlu neurons.
Previous studies indicate that SPZGABA neurons act as a necessary relay for circadian behavior. Here, the authors examined whether SPZ maintains circadian pain sensitivity in CCI day 7 mice. Retrograde AAV-Vgat-Cre was injected into PVN and AAV-DIO-jGCaMP8m into SPZ of C57BL/6J mice, and fiber photometry recordings showed that Ca²⁺ event frequency and peak in SPZ neurons projecting to PVN were higher at ZT14 than ZT5 in both sham and CCI mice (Figures 4O-Q), synchronized inversely with SCN activity and pain sensitivity.
To test necessity, Vgat-Cre mice received retrograde AAV-DIO-Flp in PVN, AAV-fDIO-Caspase3 in SPZ, and AAV-DIO-jGCaMP8m in vlPAG (Figure 4R). Fiber photometry showed that ablation of SPZ neurons projecting to PVN reduced vlPAGGABA Ca²⁺ activity at ZT5 below ZT14 (Figures 4S-T). Behavioral testing revealed a reversal of circadian pain sensitivity, with ZT14 thresholds lower than ZT5 (Figures 4U-V), confirming that SPZ neurons are essential for maintaining circadian pain sensitivity and ensuring higher daytime pain in CCI mice.
Chemogenetic modulation of the SCN → SPZ → PVN circuit in CCI day 7 mice (retrograde AAV-Vgat-Cre in PVN, AAV1-hSyn-Flp in SCN, and dual-dependent AAV in SPZ) showed that activation at ZT5 increased pain thresholds, whereas inhibition at ZT14 or local SPZGABA neuron inhibition reduced pain thresholds. Overall, SPZ functions as an inhibitory relay, and artificial modulation can reverse the circadian rhythm of pain sensitivity in neuropathic pain mice.
Figure 4. SPZ plays a key role in maintaining daily fluctuations of pain sensitivity
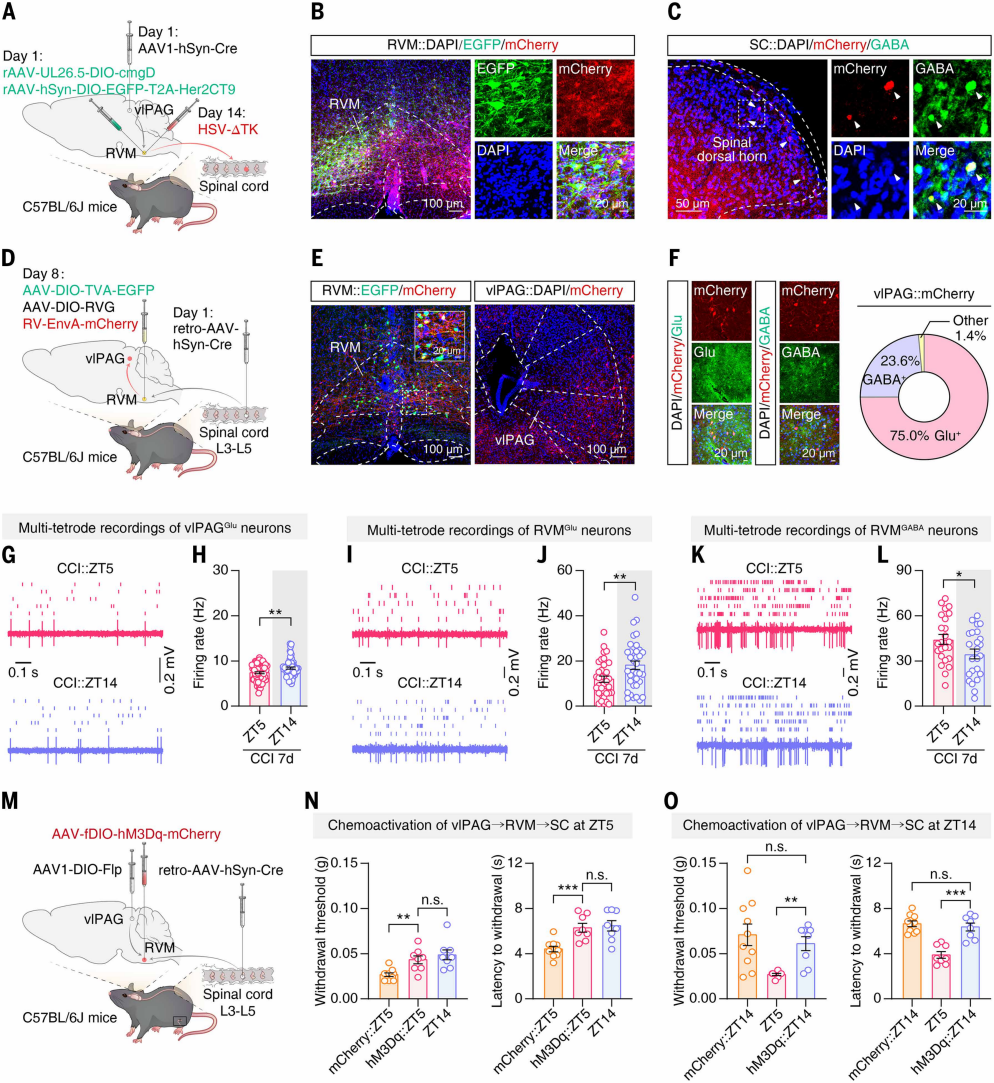
vlPAGGABA neurons are key components of the descending pain modulatory network. The authors investigated whether their rhythmic activity regulates circadian pain sensitivity. Using Cre-dependent anterograde trans-synaptic tracing, AAV1-hSyn-Cre was injected into vlPAG of C57BL/6J mice, and helper virus was injected into RVM (Figures 5A-B). Two weeks later, H129-ΔTK virus was injected at the same RVM site. After viral expression, mCherry⁺ neurons co-labeled with GABA antibodies were detected in the spinal dorsal horn (Figure 5C), indicating that RVM neurons innervated by vlPAG project to GABAergic inhibitory neurons in the dorsal horn. Retrograde tracing was performed by injecting AAV-hSyn-Cre into the L3-L5 dorsal horn and helper virus plus RV-ΔG-mCherry into RVM (Figures 5D-E). Three weeks later, mCherry⁺ neurons in vlPAG were primarily glutamatergic, with a minority GABAergic (Figure 5F), showing that vlPAG^Glu neurons provide the main input to RVM neurons projecting to the spinal cord.
Given the rhythmic activity of vlPAGGABA neurons, their effect on circuit activity was examined. In vivo multi-electrode recordings revealed that in CCI day 7 mice, vlPAGGlu neuron activity at ZT5 was lower than at ZT14 (Figures 5G-H), opposite to vlPAGGABA activity (ZT5 higher than ZT14); sham controls showed no difference. This suggests that the rhythmic activity of vlPAGGlu neurons is regulated by inhibitory input from vlPAGGABA neurons. Multi-electrode recordings also showed that RVMGlu neuron firing was lower at ZT5 than ZT14 (Figures 5I-J), whereas RVMGABA neuron firing was higher at ZT5 than ZT14 (Figures 5K-L); sham controls showed no difference, confirming circadian rhythmic activity in RVM neurons of CCI mice.
To test whether circuit rhythmicity regulates pain sensitivity, vlPAG of mice was injected with AAV1-DIO-Flp, L3-L5 dorsal horn with retrograde AAV-hSyn-Cre, and RVM with the corresponding virus (Figure 5M). Activation of the circuit at ZT5 increased CCI mouse pain thresholds to near ZT14 levels (Figure 5N), whereas inhibition at ZT14 decreased thresholds to ZT5 levels. Activation at ZT14 or inhibition at ZT5 had no effect (Figure 5O), indicating that modulation of this circuit can disrupt circadian pain sensitivity.
Figure 5. Rhythmic activity of the vlPAG → RVM → spinal cord circuit regulates circadian pain sensitivity
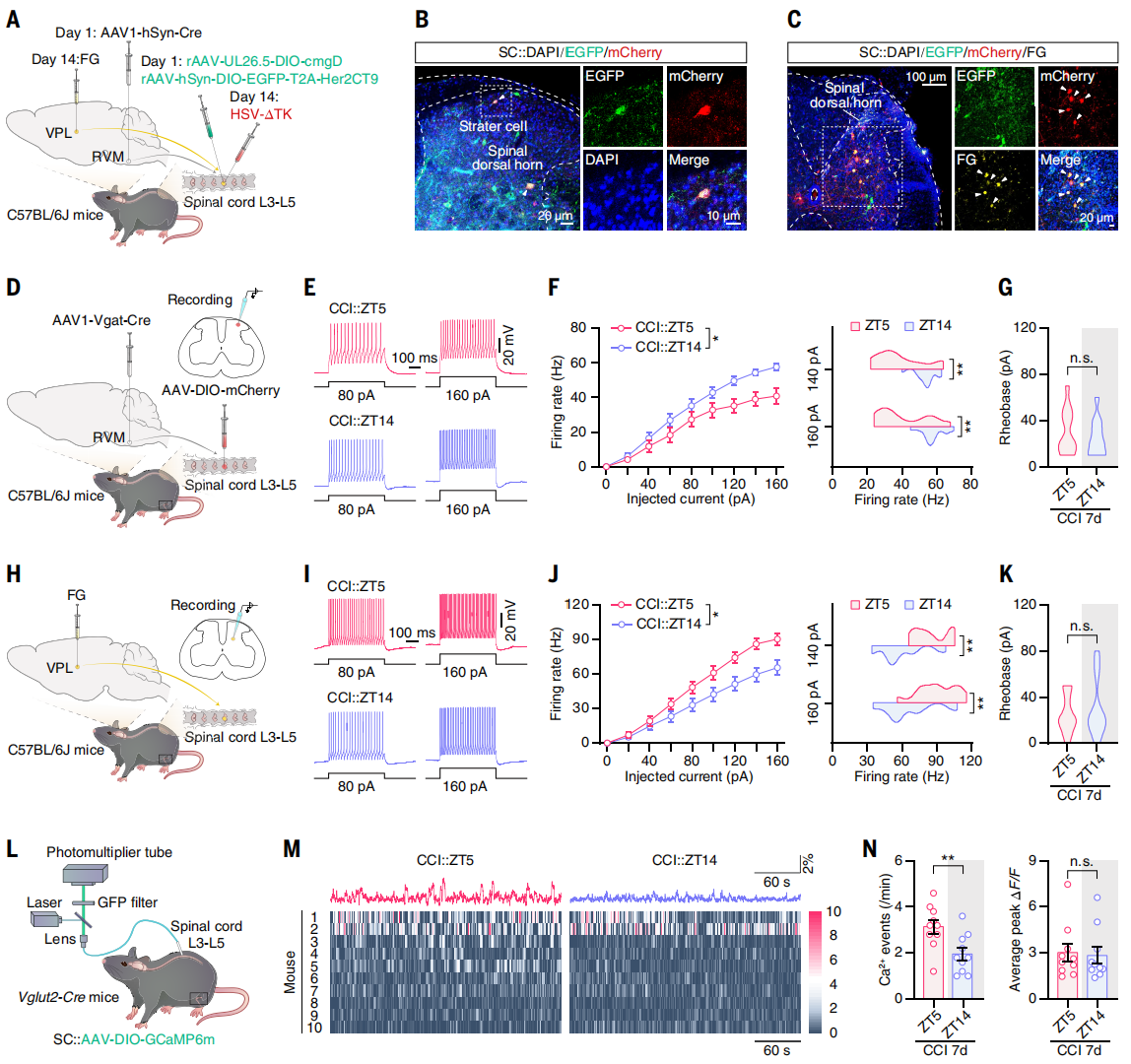
Rhythmic Activity of Spinal Cord GABA (SCGABA) Neurons Drives Circadian Pain via Upstream SCGlu Neurons
The authors investigated whether the descending vlPAG → RVM → spinal GABAergic (SCGABA) circuit regulates spinal ascending projection neurons. The ventral posterolateral thalamus (VPL) is a classical relay for the spinothalamic tract. Retrograde tracer FG was injected into VPL, and one week later, FG⁺ cells were detected in the spinal cord, ~81.1% of which co-expressed glutamate.
To determine whether SCGABA neurons receiving RVM input project to VPL-related SCGlu neurons, combined anterograde and retrograde tracing was performed: AAV1-hSyn-Cre was injected into RVM, and helper virus into the spinal cord, followed by H129-ΔTK virus and FG. EGFP⁺/mCherry⁺ co-labeled cells were identified as RVM-origin starter cells (Figure 6B), and FG⁺ cells (VPL retrograde-labeled) co-localized with mCherry⁺ cells (Figure 6C), confirming direct synaptic connectivity.
To test rhythmicity, patch-clamp recordings were performed on spinal cord slices (Figure 6D). In CCI day 7 mice, SCGABA neurons receiving RVM input fired fewer action potentials at ZT5 than ZT14, while baseline currents were unchanged (Figures 6E-G). Fiber photometry confirmed lower Ca²⁺ activity at ZT5 than ZT14; sham controls showed no difference, demonstrating circadian rhythmicity.
Additionally, VPL retrograde-labeled FG⁺ neurons showed higher excitability at ZT5 than ZT14 (Figures 6H-K), and SCGlu neuron Ca²⁺ activity was higher at ZT5 than ZT14 (Figures 6L-N); sham controls showed no difference. Since SCGABA neurons can regulate spinal projection neurons, the phase-opposite rhythm of SCGlu neurons likely arises from inhibitory input.
Figure 6. Rhythmic SCGABA neuron activity synchronizes daily pain sensitivity via modulation of ascending SCGlu neurons
Summary
Neuropathic and inflammatory pain in mice exhibits intrinsic circadian fluctuations in pain sensitivity, regulated by the hypothalamic suprachiasmatic nucleus (SCN) as the master clock, with SCNVIP neurons as the central node. Circadian pain is driven by the multi-synaptic SCNVIP → SPZGABA → PVNGlu → vlPAGGABA circuit and executed via the descending vlPAG → RVM → spinal cord analgesic system, with SPZ acting as an essential inhibitory relay between SCN and PVN.
All the viral tools used in this study are available from Brain Case Biotech
Brain Case Biotech also offers various custom services—please contact bd@ebraincase.com for more information.
Service Type :
Select the service you'd like to purchase.
Order Information(Premade-AAVs)
Please provide us some information about the service you'd like to order.
Order Information(Custom AAV/Lentivirus)
Please provide us some information about the service you'd like to order.
Order Information(Others)
Please provide us some information about the service you'd like to order.