Nature Methods | CaBLAM: A Novel Long-Term Bioluminescent Calcium Indicator Without Excitation Light
Release time:2026-05-07 15:36:09
Calcium imaging is a core technique for monitoring intracellular calcium levels and understanding signal transduction across cell types and whole organisms, with widespread applications in systems neuroscience. Current fluorescent genetically encoded calcium indicators (GECIs), such as GCaMP8 and the GECO family, require high-intensity excitation light, which can cause photobleaching, background autofluorescence, and phototoxicity, limiting long-term imaging. Bioluminescent GECIs, on the other hand, do not require excitation light but suffer from low dynamic range and calcium affinity that does not match physiological levels, making them suitable only for population-level recordings.
On December 2, 2025, the Shaner Laboratory at the University of California, San Diego, published a study in Nature Methods titled “CaBLAM: a high-contrast bioluminescent Ca²⁺ indicator derived from an engineered Oplophorus gracilirostris luciferase.” The paper introduces CaBLAM, a high-contrast bioluminescent calcium indicator developed from a shrimp luciferase, achieving ~83-fold signal contrast in vitro and a calcium affinity (KD ≈ 439 nM) compatible with physiological levels. By eliminating the need for excitation light, CaBLAM avoids photobleaching and phototoxicity. In cultured neurons, CaBLAM enables single-cell calcium imaging at 10 Hz, and in awake mice and zebrafish, it supports hours-long continuous imaging, with single-trial signal-to-noise ratios exceeding those of GCaMP6s.
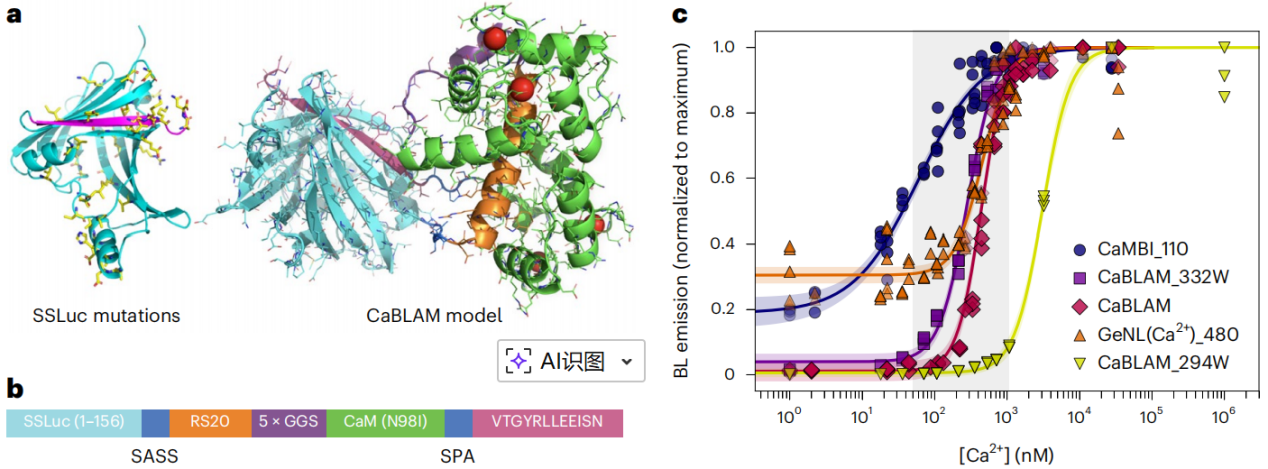
CaBLAM is a high-contrast bioluminescent calcium indicator developed from a rationally engineered shrimp luciferase (OLuc). Through directed evolution of the OLuc mutant eKAZ, the researchers obtained a highly active and soluble luciferase scaffold, SSLuc (Fig. 1a). In vitro, when using the substrate furimazine (Fz), SSLuc exhibited higher activity than NanoLuc, while remaining resistant to aggregation and compatible with domain insertions.
Next, the last two β-strand connecting loops of SSLuc were split to insert the calcium-sensing domain RS20–CaM (Fig. 1a, b), retaining the N-terminal mNeonGreen to enhance bioluminescence via FRET, while restricting signal readout to the bioluminescence channel to avoid fluorescence interference.
Early clones had excessively high calcium affinity (KD < 10 nM), so the RS20/CaM sequence from GCaMP6s and the CaM mutant N98I were used to tune the apparent KD to ≈439 nM, matching physiological calcium levels in hippocampal neurons. The C-terminal peptide was further optimized to VTGYRLLEEISN, replacing lysine with glutamate to weaken peptide–enzyme interaction, achieving an in vitro contrast of 83-fold (Fig. 1c).
Additionally, two variants were created to cover different cellular calcium ranges: 294W (N98W in CaM; KD ≈ 3 μM) and 332W (Q135W in CaM; KD ≈ 280 nM). The final CaBLAM inherits the high activity and stability of SSLuc, addressing the low dynamic range and mismatched affinity of traditional bioluminescent indicators, making it a versatile tool for long-term, high-resolution calcium imaging without excitation light.
Figure 1: CaBLAM structure and Ca²⁺ affinity.
Comparative Study of CaBLAM and GCaMP8s in Rat Hippocampal Neurons
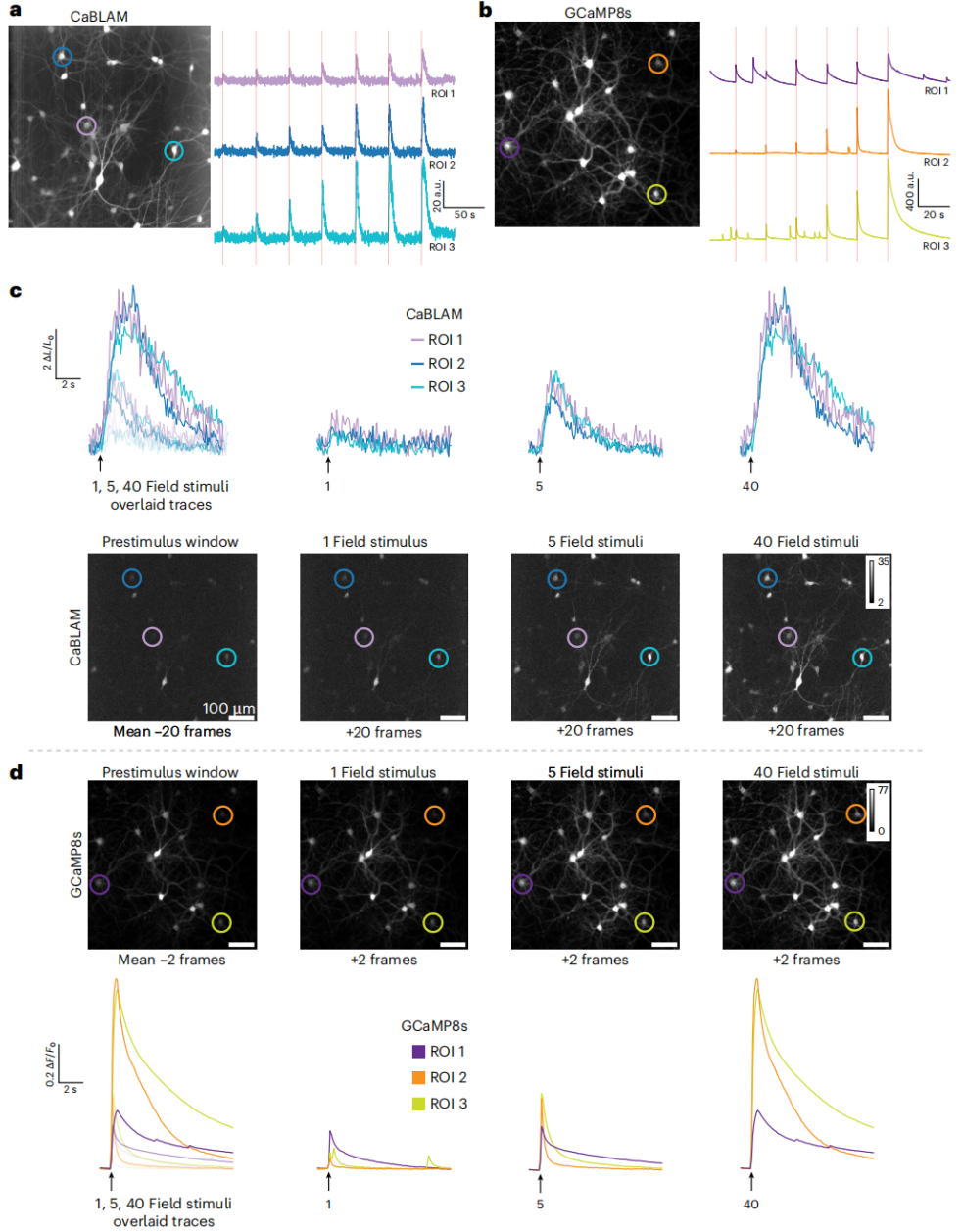
CaBLAM was characterized with synchronous calcium imaging and field stimulation, and benchmarked against the fluorescent sensor GCaMP8s in rat hippocampal neurons. At 10 Hz imaging, CaBLAM photon counts were lower, but detectable evoked responses were generated across a series of field stimulations (Fig. 2a, c). GCaMP8s exhibited higher brightness, capturing both spontaneous and evoked neuronal activity (Fig. 2b, d).
CaBLAM reliably reported intracellular calcium changes, with normalized signals (ΔL/L)—a common metric in bioluminescence imaging—showing significant fluctuations (Fig. 2c). GCaMP8s, due to high baseline fluorescence, exhibited smaller normalized fluorescence changes (ΔF/F) (Fig. 2d), but still showed observable responses, consistent with previous reports.
Figure 2: Field stimulation responses in rat hippocampal neurons expressing CaBLAM or GCaMP8s.
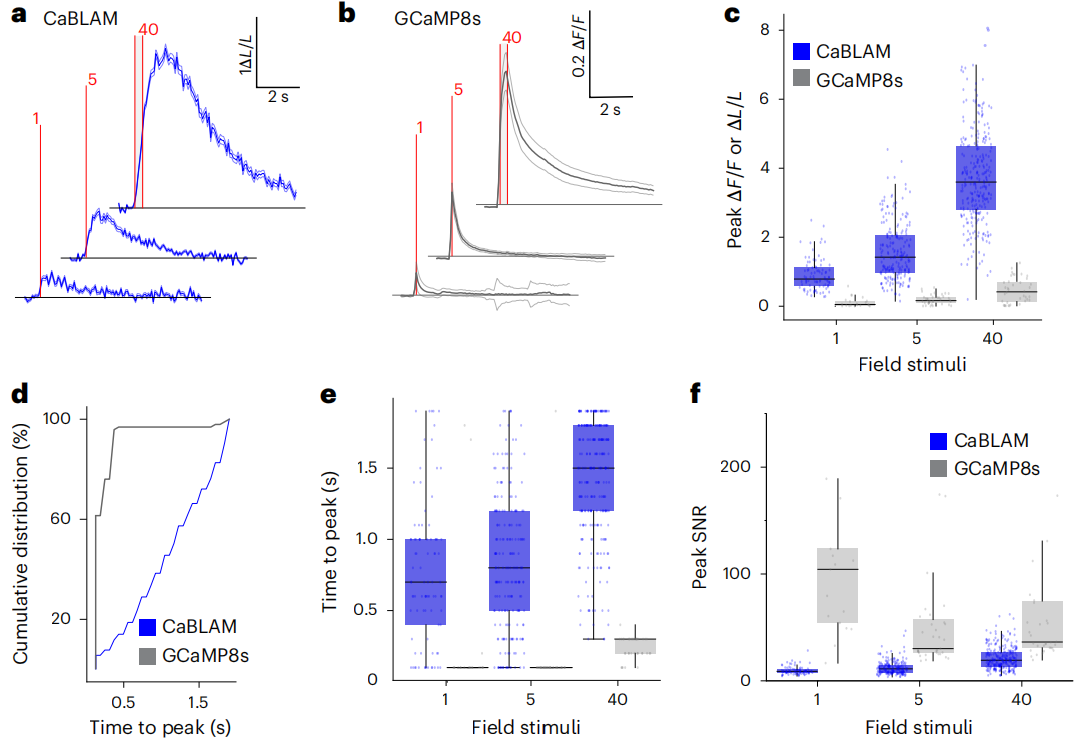
Further characterization of evoked signals in neurons expressing CaBLAM (Fig. 3a) and GCaMP8s (Fig. 3b) confirmed preliminary observations. Due to CaBLAM’s low baseline signal, ΔL/L changes under field stimulation (1, 5, 40 pulses, 83 Hz, 1 ms) were significantly higher than GCaMP8s ΔF/F (Fig. 3c). As expected, the unoptimized fast-response CaBLAM kinetics were significantly slower than GCaMP8s (Fig. 3d).
The peak response timing differed markedly: GCaMP8s peaked at ~100 ms for 1–5 pulses, extending to 300 ms under high-frequency stimulation (Fig. 3e). With 9.2 μM Fz perfusion, CaBLAM median peak times were 700, 800, and 1500 ms for 1, 5, and 40 pulses, respectively. Without synaptic blockers, true kinetics may be faster.
In terms of peak signal-to-noise ratio (SNR), GCaMP8s had higher SNR in single-layer neuronal imaging despite lower contrast (Fig. 3f). At 4.6 μM Fz, CaBLAM responses were larger and faster, while 9.2 μM Fz achieved a proportion of responsive neurons comparable to GCaMP8s, making it the optimal detection concentration.
Figure 3: CaBLAM provides high-contrast reporting of stimulation-evoked neuronal activity in cultured neurons.
In Vivo Comparison of CaBLAM and GCaMP6s
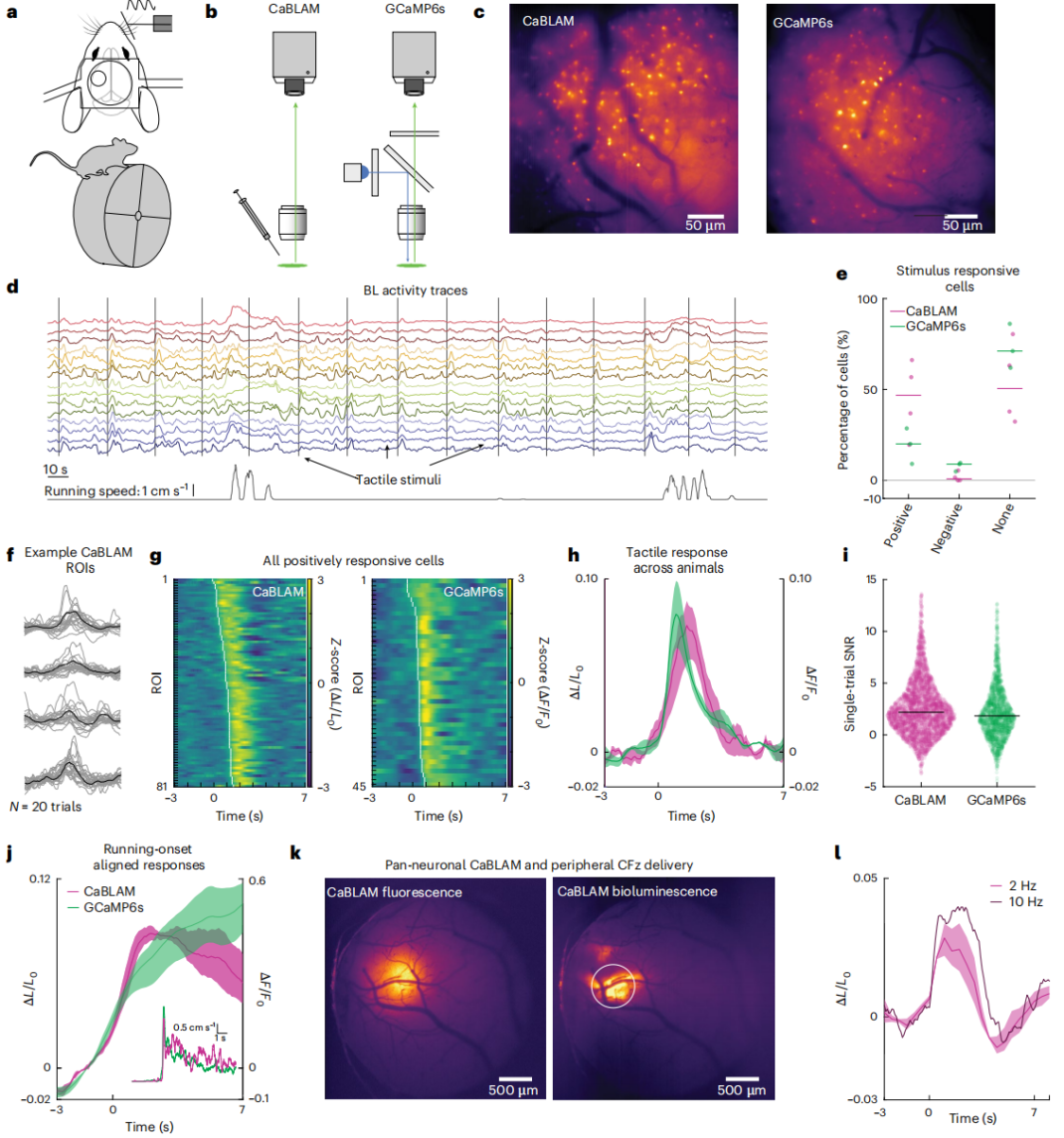
After determining fluorofurimazine (FFz, Promega, cat. no. N4110) as the optimal substrate for CaBLAM, NDNF-Cre mice cortical interneurons were used as a model (Fig. 4) to compare in vivo performance of CaBLAM and GCaMP6s. The CaBLAM group received no external light, while the GCaMP6s group was imaged with single-photon epifluorescence. Mice were head-fixed but able to run freely on a wheel, and received whisker tactile stimulation (Fig. 4a).
In the CaBLAM group, FFz was delivered via a cranial-window edge cannula (Fig. 4b, c) to directly assess single-cell evoked signals. Single-cell activity was detected in all mice (Fig. 4d). Some NDNF neurons showed positive or negative responses to tactile stimuli, modulated by locomotion/arousal state (Fig. 4e, g–j). Most responsive cells in both groups were positively modulated (Fig. 4f), and this study focused on these cells.
Tactile-evoked waveforms were highly similar between groups, with ΔF/F₀ (GCaMP6s) and ΔL/L₀ (CaBLAM) amplitudes and temporal dynamics closely matched (Fig. 4i). Using half-peak measurements, CaBLAM exhibited longer latency than GCaMP6s, consistent with in vitro experiments. Single-trial SNR was significantly higher for CaBLAM. Peak signal amplitude showed no statistical advantage, but CaBLAM’s lower baseline standard deviation underlies its superior SNR.
Peripheral injection validation: CaBLAM-expressing mice received 200 μl cephalofurimazine (CFz, Promega, cat. no. CS3553A01) into the orbit. Wide-field imaging at 2 Hz detected rapid tactile-evoked responses with latency ≤0.5 s, avoiding hemodynamic interference (Fig. 4k, l). Responses were similar at 10 Hz and 2 Hz, with no significant SNR difference. However, this method produced weaker signals in NDNF-Cre mice and could not capture calcium fluctuations.
Figure 4: CaBLAM in vivo imaging SNR is comparable to GCaMP under epifluorescence illumination.
Long-Term Imaging Validation
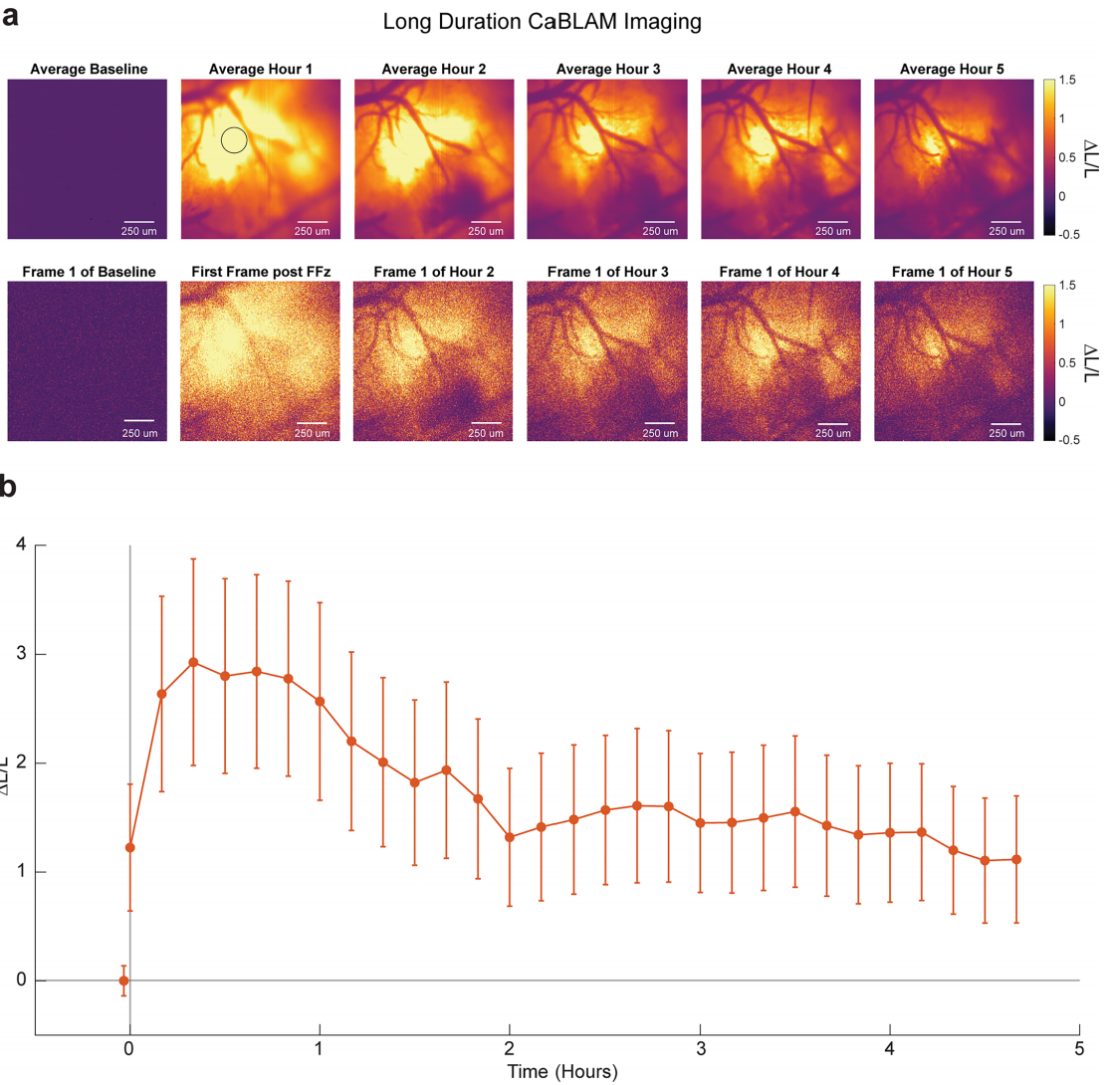
FFz was applied to the cortex of anesthetized mice, with 10 Hz imaging for over 5 hours (Fig. 5a). Signals appeared within 2 minutes, peaked at 30 minutes, then slowly declined over 2 hours, stabilizing at ~50% of peak between 3–5 hours (Fig. 5b). Responses remained stable throughout, and SNR showed no variation across time points. Even after a single FFz application for over 5 hours, signals remained robust, demonstrating CaBLAM’s suitability for long-term imaging, with duration limited by substrate bioavailability.
After imaging, neurites remained clearly visible at 40× magnification and 1 Hz acquisition. Additionally, CaBLAM bioluminescence can be used to monitor calcium dynamics in zebrafish larvae neurons and astrocytes.
Figure 5: Time course of CaBLAM bioluminescence after single FFz cortical administration in mice.
Core Advantages and Application Differences of CaBLAM Compared with Traditional Fluorescent Calcium Indicators (e.g., GCaMP Series)
Core Advantages:
1. No excitation light required, completely avoiding photobleaching, phototoxicity, and background autofluorescence.
2. Low baseline signal, yielding higher single-trial in vivo SNR than GCaMP6s.
3. Supports long-term imaging (>5 hours), whereas fluorescent indicators are typically limited to ~30 minutes due to phototoxicity.
Application Differences:
CaBLAM is ideal for long-term monitoring of neuronal activity (e.g., learning and memory), imaging in freely behaving animals, and photosensitive samples (e.g., embryonic cells).
GCaMP series is better suited for experiments requiring fast response times (peak ~100 ms vs. CaBLAM ~700 ms) and higher spatial resolution for rapid dynamic imaging.
Service Type :
Select the service you'd like to purchase.
Order Information(Premade-AAVs)
Please provide us some information about the service you'd like to order.
Order Information(Custom AAV/Lentivirus)
Please provide us some information about the service you'd like to order.
Order Information(Others)
Please provide us some information about the service you'd like to order.