- E-mail:BD@ebraincase.com
- Tel:+8618971215294
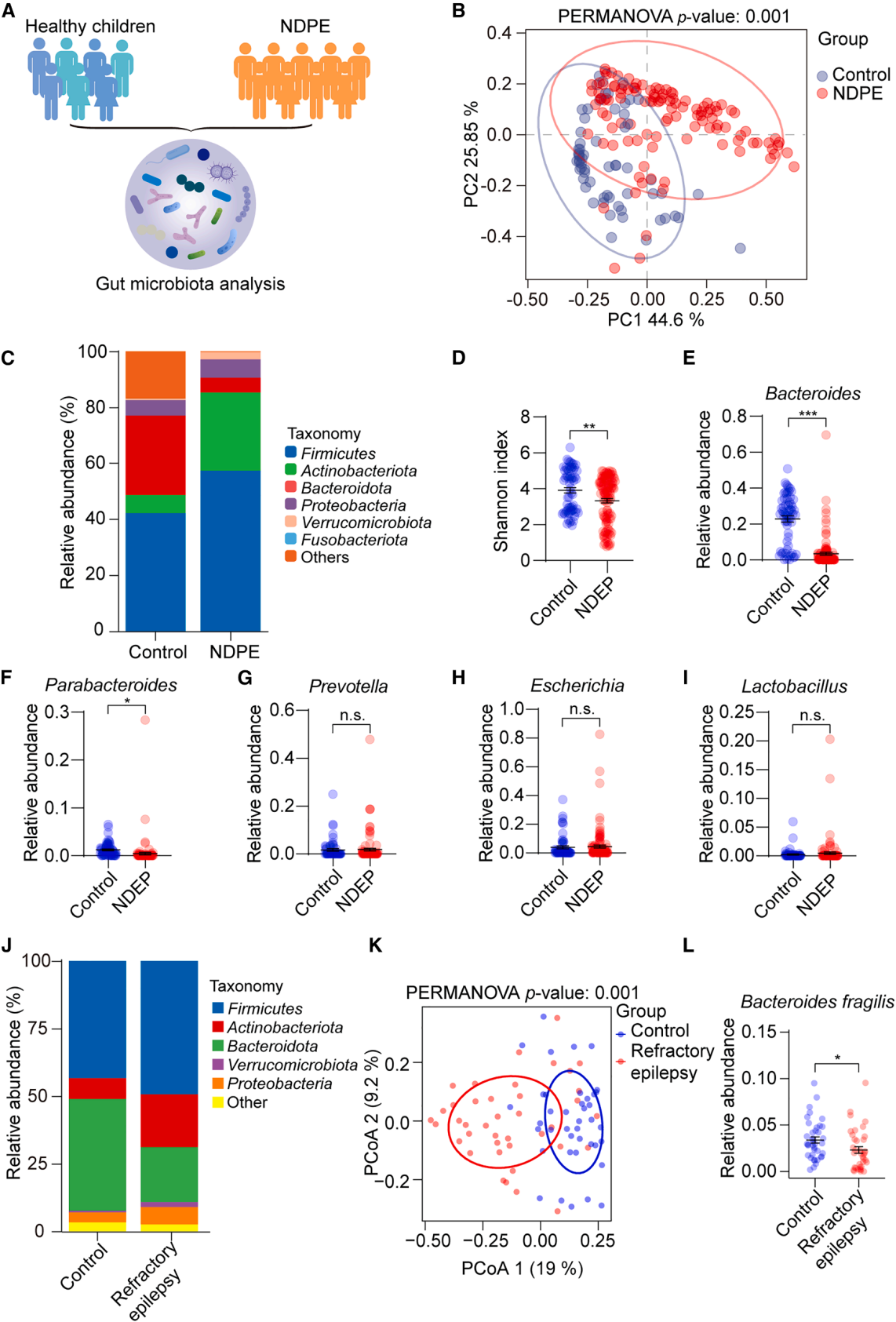
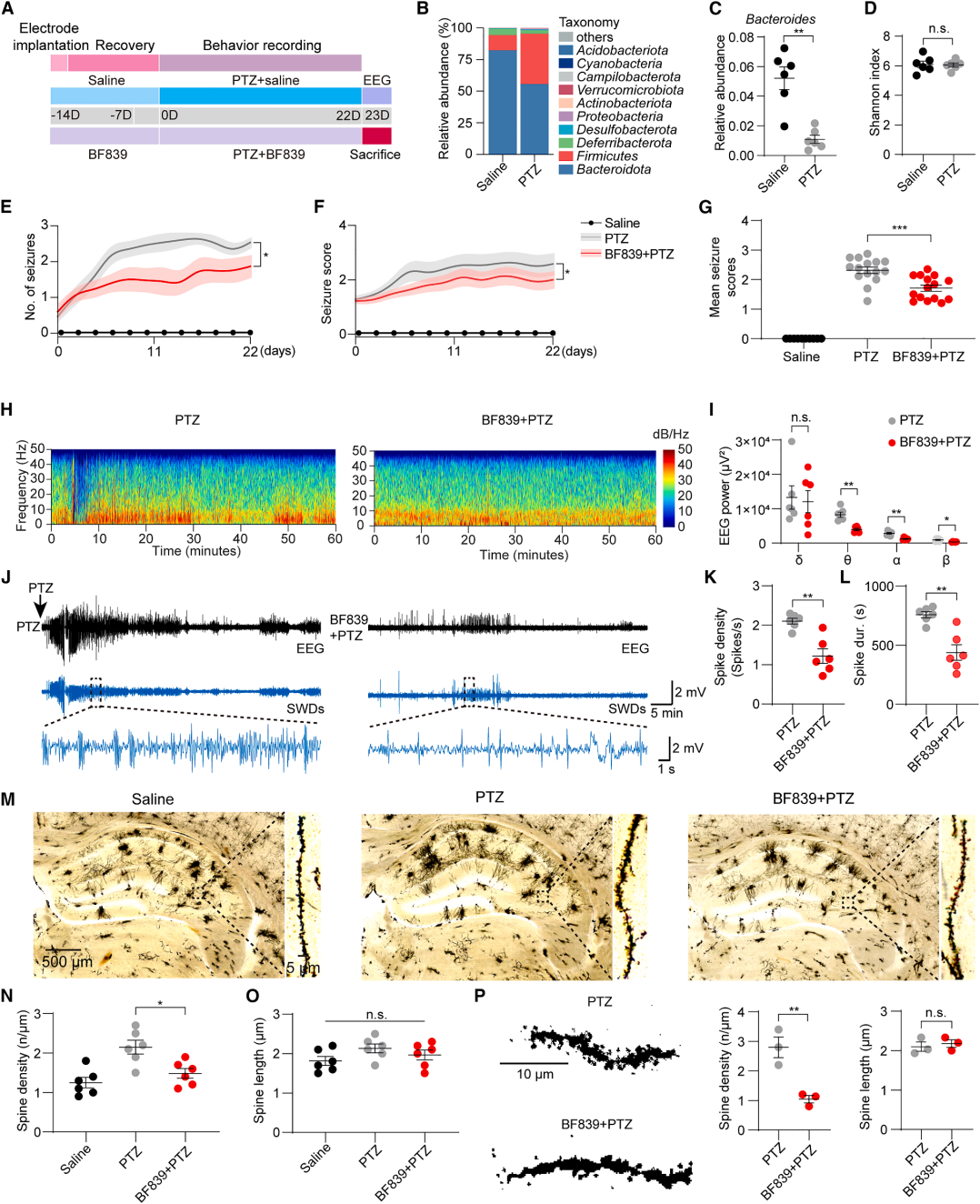
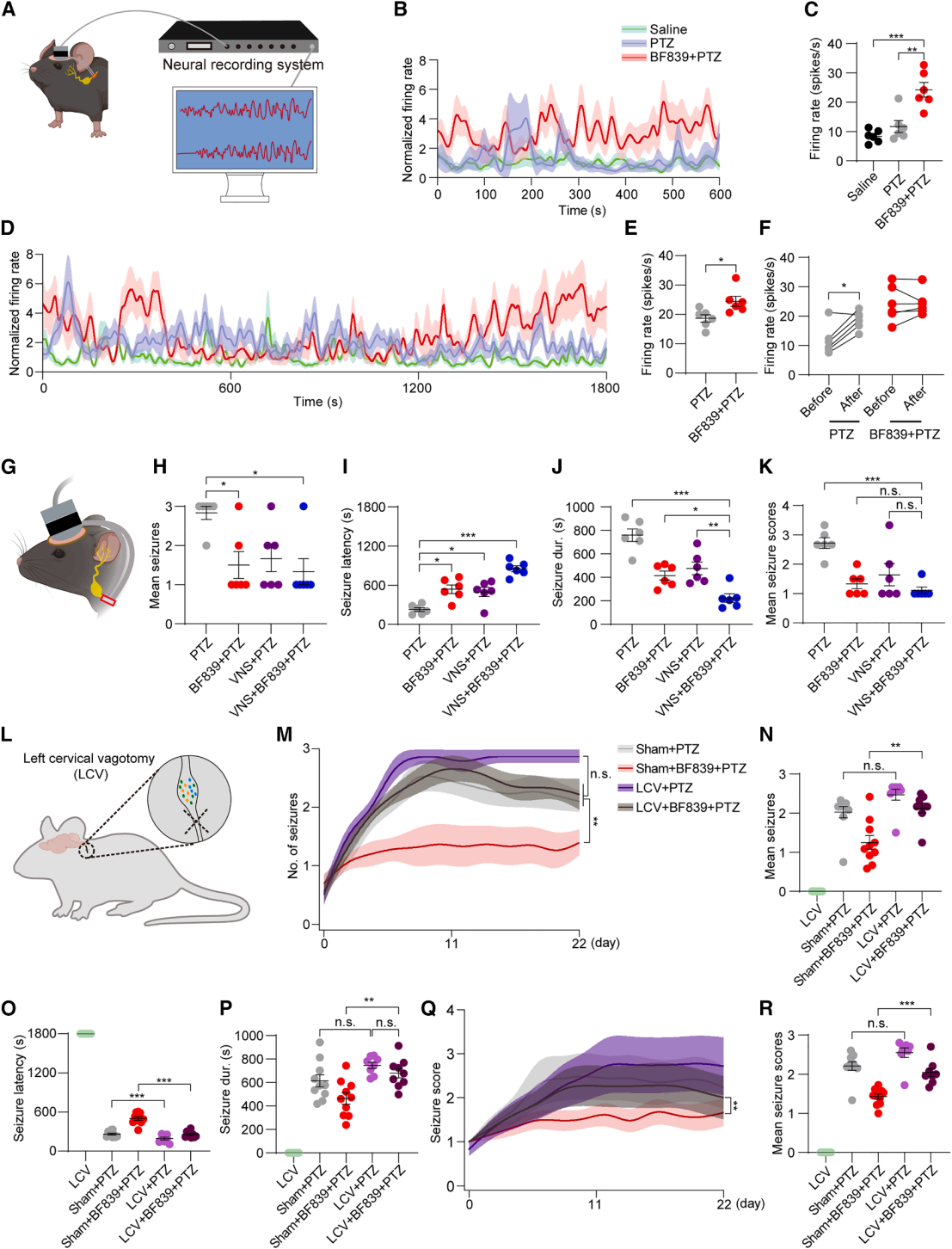
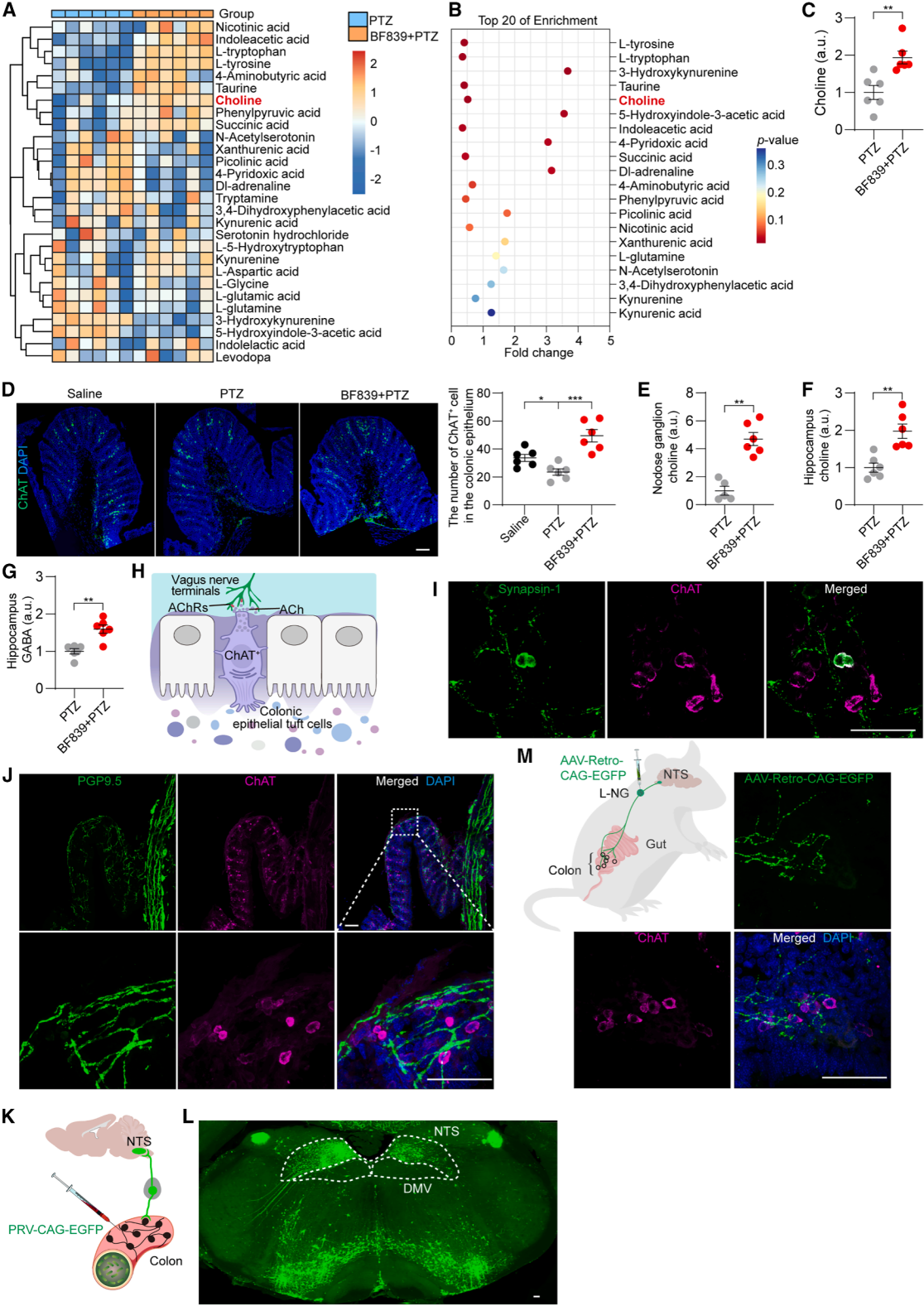
| Product Category | Product No. | Product Name |
|---|---|---|
| Sparse Labeling | BC-SL001 | NCSP-YFP-2E5 |
| Fluorescent Protein | BC-0251 | AAV9-Retro-CAG-EGFP |
| Fluorescent Protein | BC-0424 | rAAV-ChAT-DIO-EGFP |
| Chemogenetics | BC-4404 | rAAV-ChAT-DIO-hM3D(Gq)-P2A-EGFP |
| Apoptosis | BC-4405 | rAAV-ChAT-DIO-taCasp3-T2A-TEVp-P2A-EGFP |
| PRV | BC-PRV-531-Pro | PRV-CAG-EGFP |
| RV-helper | BC-2042 | rAAV-hSyn-H2B-EGFP-T2A-TVA |
| RV-helper | BC-4406 | rAAV-hSyn-N2cG |
| RV | BC-RV-CVS EnvA472 | CVS-EnvA-ΔG-mCherry-P2A-Cre |


WhatsApp Business Account
Address:-
Address:-



