Customer Article | Cell | Fudan University’s YiZhou Jiang, ZhiMin Shao, JinFei Ni, and Yi Xiao Team Reveal the Role of Sensory Neurons in Regulating the Tumor Microenvironment in Triple-Negative Breast Cancer (TNBC),Providing New Targets for Immunotherap
Release time:2026-02-28 13:51:03
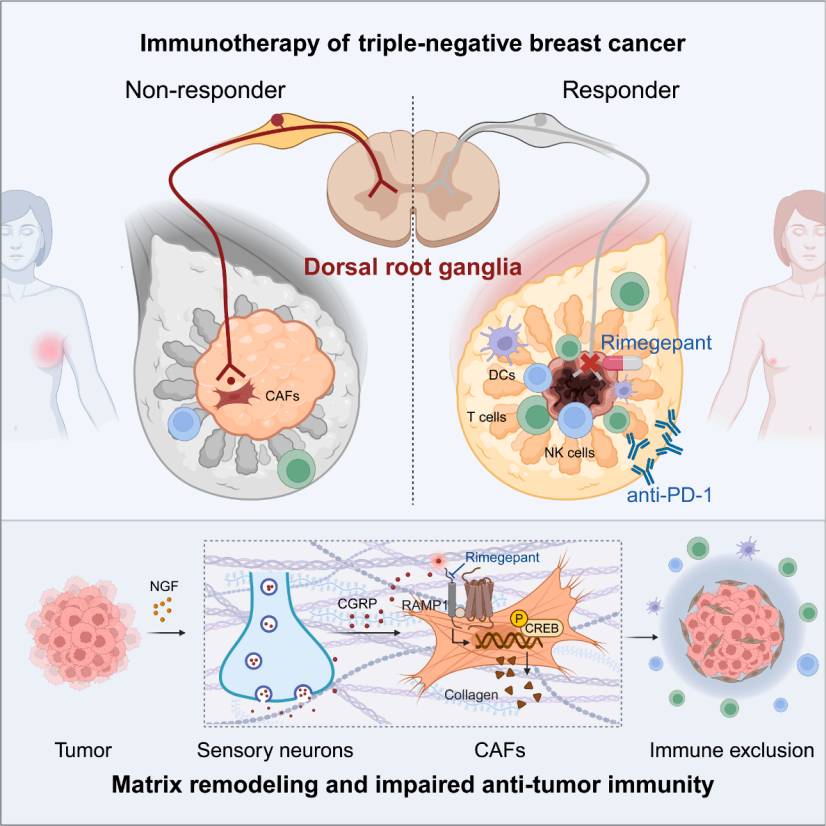
Triple-negative breast cancer (TNBC), which lacks estrogen receptors, progesterone receptors, and HER2 expression, is the most invasive subtype of breast cancer. Immune checkpoint blockade (ICB) therapy, which works by blocking key immune checkpoints such as CTLA-4, PD-1/PD-L1 to relieve the tumor microenvironment's suppression of T cells and restore the immune system's ability to recognize and kill tumor cells, holds potential. However, only a subset of patients benefit, and tumor microenvironment (TME) dysregulation is a major cause of resistance. Neuroinnervation plays a critical role in tumor progression, but the role of sensory neurons in the TNBC ecosystem remains unclear.
On February 6, 2026, a team from the Fudan University Shanghai Cancer Center led by YiZhou Jiang, ZhiMin Shao, and Yi Xiao, in collaboration with the team of JinFei Ni from the Fudan University Institute of Neuroscience, published a paper titled “Sensory neurons drive immune exclusion by stimulating a dense extracellular matrix in the breast cancer tumor microenvironment” in the prestigious journal Cell. This study reveals a regulatory axis involving "tumor cells - nerve growth factor (NGF) - sensory neurons - calcitonin gene-related peptide (CGRP) - cancer-associated fibroblasts (CAFs) - collagen," elucidating the molecular mechanism by which sensory neurons drive immune exclusion in TNBC. The study confirms that the CGRP receptor RAMP1 (receptor activity-modifying protein 1) antagonist, rimegepant, can act as an immune sensitizer and, when combined with anti-PD-1 therapy, can enhance the treatment of TNBC, providing a basis for clinical drug repositioning.
Perineural Infiltration (PNI) Is Associated with Dysregulated Tumor Microenvironment (TME) and Dominated by Sensory Neurons in TNBC
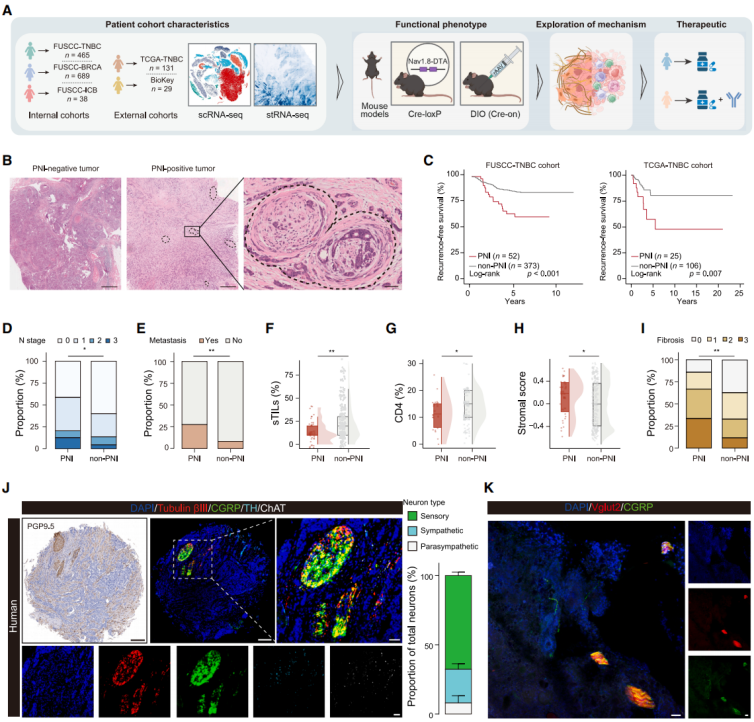
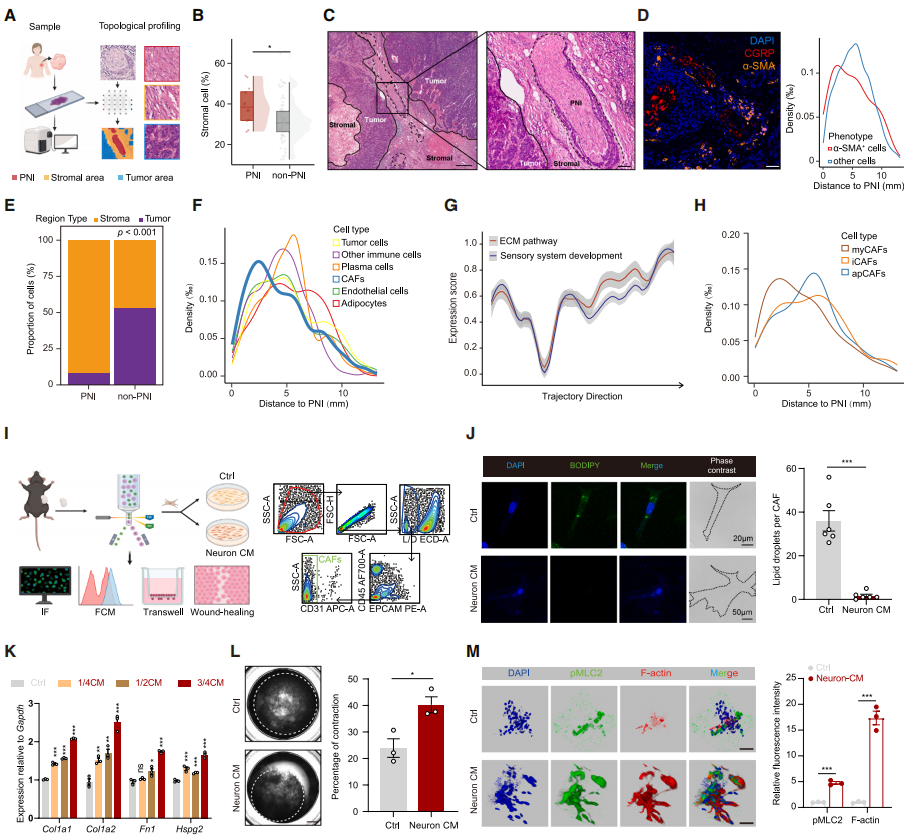
To investigate the role of perineural infiltration (PNI) in TNBC, multi-omics data and mouse models were analyzed. The Fudan University Shanghai Cancer Center (FUSCC)-TNBC cohort and The Cancer Genome Atlas (TCGA) cohort showed that PNI-positive patients had shorter recurrence-free survival and were associated with advanced lymph node staging and high bone metastasis rates (Fig. 1A-E). Multivariate Cox analysis confirmed that PNI is an independent poor prognostic factor for TNBC. PNI is unrelated to baseline clinical pathological features but significantly affects the TME: it is negatively correlated with immune infiltration, positively correlated with CAF abundance, and reduces tumor-infiltrating lymphocytes (TILs), CD4⁺ T cells, and tertiary lymphoid structures (TLSs)/lymphoid aggregates (LAs) (Fig. 1F, 1G), while promoting stroma activation and fibrosis (Fig. 1H, 1I), indicating that PNI may worsen patient survival by disrupting the TME. Multiplex immunofluorescence showed that sensory neurons account for about 70% of the nerve fibers in TNBC (Fig. 1J, 1K); CTB retrograde labeling confirmed their origin from T11-L4 dorsal root ganglia (DRGs). Furthermore, PNI has prognostic significance only in TNBC and not in hormone receptor (HR)⁺/HER2⁻ or HER2⁺ breast cancer. In conclusion, PNI is a poor prognostic factor in TNBC, associated with fibrosis-promoting and immune-suppressive TME, with sensory neurons from the T11-L4 segment of DRGs being the main neuronal type in TNBC.
Fig. 1. PNI is related to TME dysregulation and is dominated by sensory neurons in TNBC
Sensory Neurons Accelerate Tumor Progression by Stimulating Collagen Deposition and Reducing Immune Infiltration
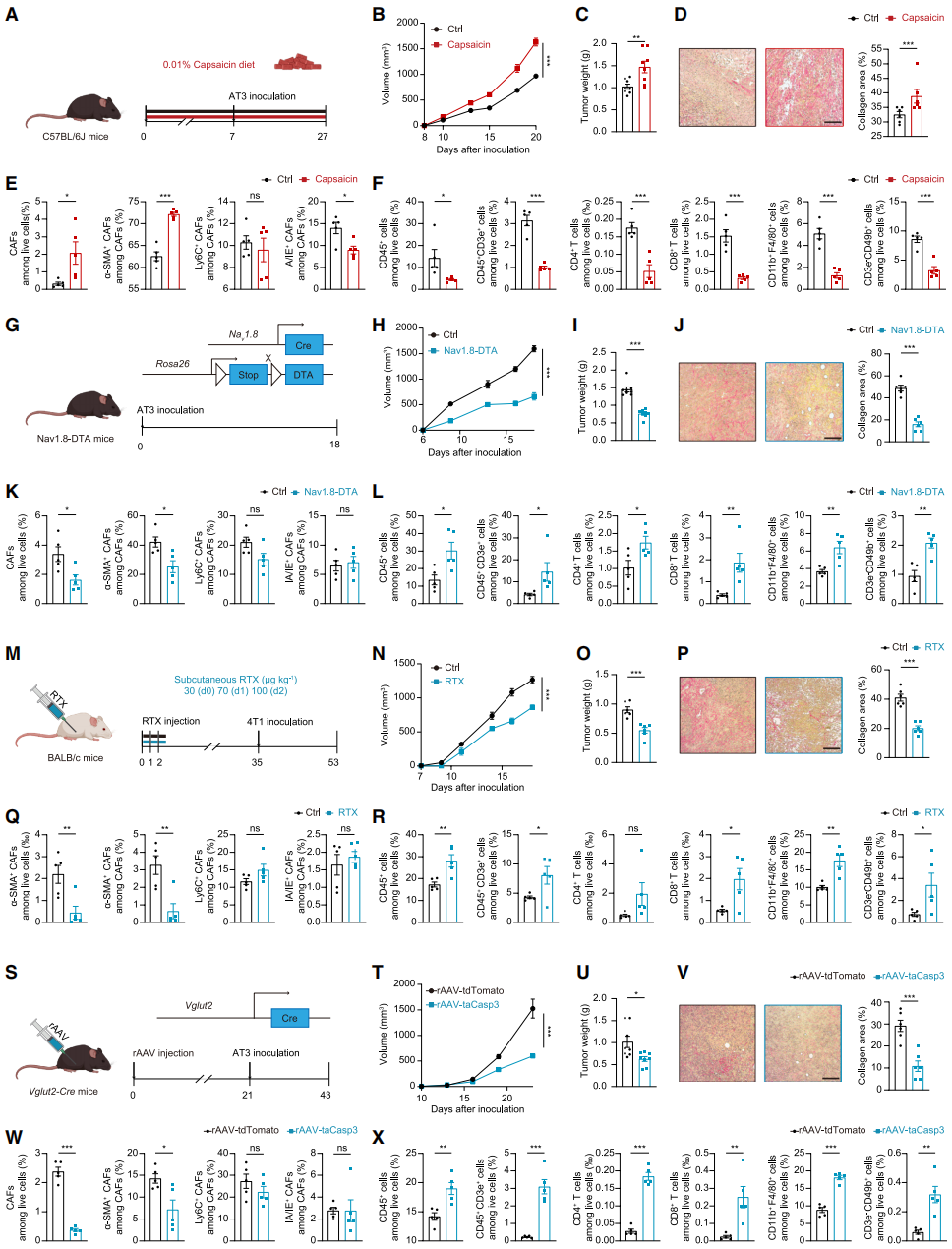
Several mouse models were used to validate the driving role of sensory neuron infiltration in promoting connective tissue proliferation and immune suppression. A low-dose capsaicin diet was used to construct a systemic sensory neuron activation model, and results showed that sensory neuron activation significantly accelerated tumor growth and fibrosis (Fig. 2A-2D). Flow cytometry confirmed that after capsaicin treatment, the proportion of CAFs in tumors increased, mainly enriching myofibroblast-like CAFs (myCAFs) (Fig. 2E). Further examination showed that sensory neuron activation suppressed the infiltration of various immune cells (Fig. 2F). In a Nav1.8-DTA systemic sensory neuron ablation model (Fig. 2G), compared with the control group, the mice showed slower tumor growth, fewer CAFs (especially myCAFs), reduced collagen deposition, improved TME, and increased infiltration of CD4⁺/CD8⁺ T immune cells (Fig. 2H-2L).
Using resin toxin (RTX), a potent capsaicin analog, to chemically ablate sensory neurons yielded similar results: tumor growth was slower, collagen was reduced, and immune cell infiltration increased (Fig. 2M-2R). To rule out systemic sensory system effects, rAAV-DIO-Caspase3 was injected into the inguinal mammary fat pad of Vglut2-Cre mice to ablate sensory neurons locally in the tumor. Despite intact systemic sensory neurons, the rAAV-treated group showed significantly slower tumor growth (Fig. 2T-2U), reduced CAFs and collagen deposition (especially myCAFs and type I collagen) (Fig. 2V-2W), and restored immune cell function, highlighting the critical role of local sensory neuron infiltration (Fig. 2X). In conclusion, sensory neurons promote the formation of myCAF phenotypes, driving fibrosis and immune evasion in TNBC.
Fig. 2. Sensory neurons accelerate tumor progression by stimulating collagen deposition and reducing immune infiltration
Tumor-Derived NGF Promotes Axonal Growth and CGRP Secretion in Sensory Neurons
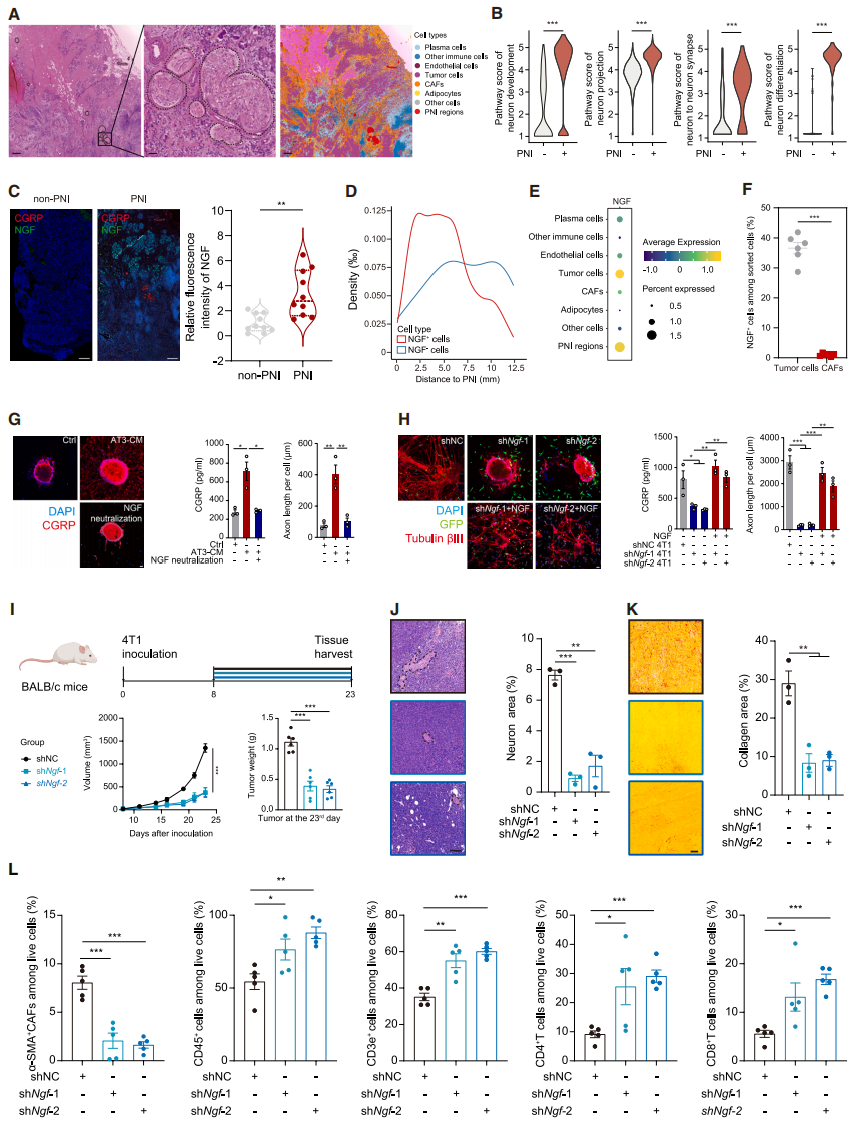
To clarify the sensitization mechanism of sensory neurons in TNBC, the researchers conducted spatial transcriptomics sequencing (stRNA-seq) at the single-cell level to identify multiple cell types and mark PNI regions (Fig. 3A). Neuron-related pathways were significantly enriched in the PNI region (Fig. 3B), and spatial trajectory analysis showed that nerve growth factor (NGF) was specifically enriched near the PNI region. mIF further confirmed that NGF levels were higher in PNI-positive samples (Fig. 3C). NGF⁺ cells were enriched near the PNI region (Fig. 3D), suggesting that NGF promotes nerve infiltration. Previous studies have shown that both cancer cells and CAFs can secrete NGF, but spatial analysis and flow cytometry confirmed that NGF primarily comes from tumor cells in TNBC. Experimental validation showed that the NGF level in AT3 tumor cell-conditioned medium (AT3 CM) increased, and sensory neurons cultured in it exhibited increased CGRP secretion and denser axonal growth, effects which could be reversed by neutralizing NGF (Fig. 3G). Knockdown of Ngf in 4T1 cells led to reduced CGRP secretion and shorter axons in cultured sensory neurons, and adding recombinant NGF reversed this effect (Fig. 3H). In vivo experiments showed that tumors grew slower in the shNgf group (Fig. 3I), the PNI area was smaller than in the shNC group (Fig. 3J), collagen deposition in the TME (especially type I) was alleviated (Fig. 3K), and immune cell infiltration increased (Fig. 3L). In conclusion, tumor-secreted NGF can attract sensory neurons, disrupt the TME, and promote tumor growth.
Fig. 3. Tumor-secreted NGF promotes axonal growth and CGRP secretion in sensory neurons
Sensory Neurons Neighbor CAFs and Promote Collagen Deposition
The researchers explored the downstream mechanisms of sensory neuron-induced collagen deposition and immune suppression using a machine learning framework based on single-cell morphology and topological mapping. Analysis of HE images from 81 TNBC patients revealed that PNI-positive patients exhibited significant accumulation of stromal cells (Fig. 4A, 4B), with PNI mainly located in the stromal area (Fig. 4C). mIF confirmed that sensory neurons (CGRP⁺) were adjacent to CAFs (α-SMA⁺) (Fig. 4D). Spatial transcriptomic analysis showed that most PNI was located in the stromal area, with CAFs enriched in the adjacent regions and immune cells excluded (Fig. 4E, 4F). These results were highly consistent with co-expression of extracellular matrix (ECM) and sensory system development pathways (Fig. 4G). In combination with the sensory neuron ablation model, which significantly reduced CAFs (Fig. 2E, 2K, 2Q, 2W), this suggests that CAFs are the target of sensory neuron action.
To clarify the effect of sensory neurons on CAFs, they divided CAFs into three subtypes: myCAFs, inflammatory CAFs (iCAFs), and antigen-presenting CAFs (apCAFs), with myCAFs located nearest to PNI (Fig. 4H). Sensory neurons were isolated from DRGs, and CAFs were sorted from AT3 tumors. After four passages in 2D culture, myCAFs stabilized at around 80%. Sensory neuron-conditioned medium (neuron CM) significantly activated CAFs (reducing lipid droplets, increasing cell volume) (Fig. 4J), increasing the proportion of myCAFs. In 3D culture, neuron CM treatment upregulated myCAF markers (Fig. 4K) and induced CAFs to produce large amounts of collagen (Fig. 4L, 4M). In conclusion, sensory neuron-driven activation of myCAFs may contribute to the formation of an immune-suppressive TME.
Fig. 4. Sensory neurons neighbor CAFs and promote collagen deposition
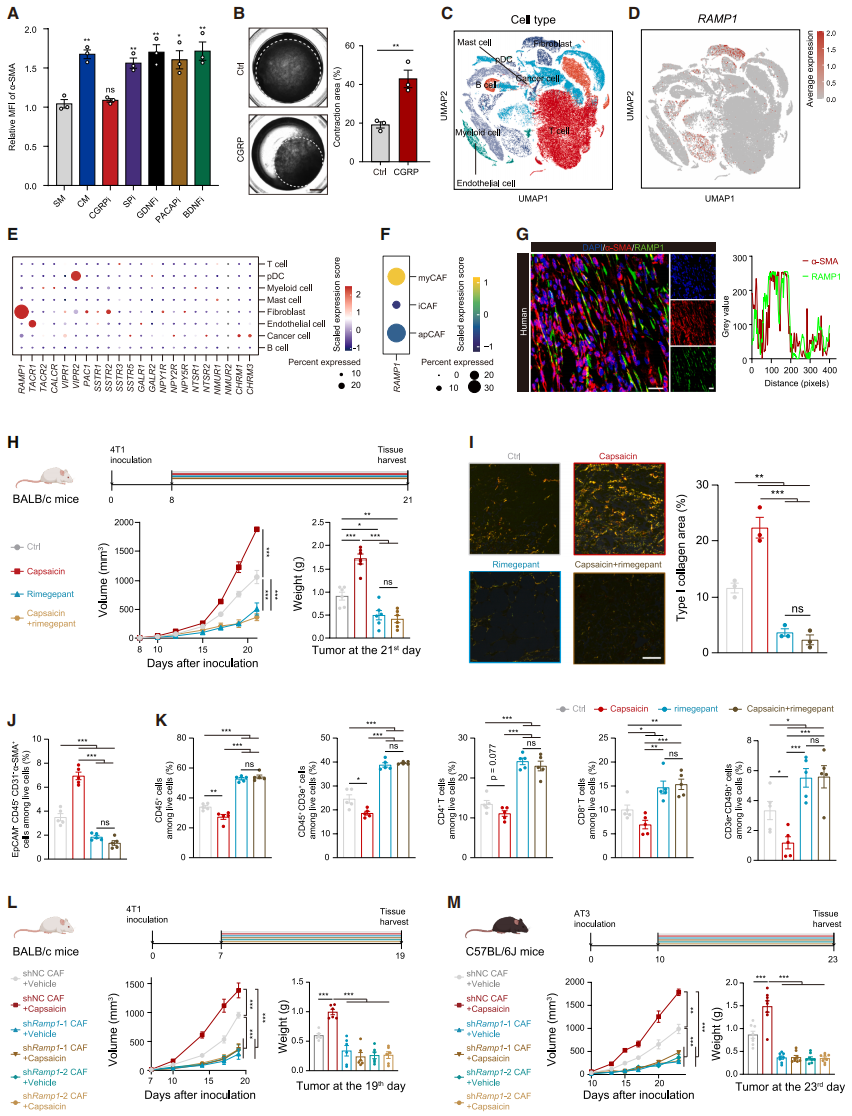
CGRP from Sensory Neurons Binds to RAMP1 on CAFs to Promote Collagen Secretion
To explore the mechanism by which sensory neurons promote myCAF activation, the researchers found that only small molecule peptides in sensory neuron-conditioned medium (neuron CM) could promote the expression of α-SMA. Among the main neuropeptide inhibitors, only the CGRP inhibitor rimegepant was able to antagonize the myCAF phenotype induced by neuron CM in CAFs (Fig. 5A). CGRP did not enhance CAF proliferation but could activate resting CAFs, increase myCAF migration, and promote collagen contraction (Fig. 5B). Single-cell RNA sequencing (scRNA-seq) data showed that only the key component of the CGRP receptor, receptor activity-modifying protein 1 (RAMP1), was highly expressed in CAFs (especially in myCAFs) (Fig. 5C-5F), and was strongly positively correlated with myCAF markers. Multiplex immunofluorescence (mIF) confirmed that RAMP1 co-localized with α-SMA in both TNBC patient samples and mouse models (Fig. 5G), suggesting that CGRP secreted by sensory neurons is a key neuropeptide driving the activation of myCAFs.
Further evaluation of the CGRP-RAMP1 axis revealed that rimegepant, which blocks RAMP1, significantly inhibited tumor growth in both normal and capsaicin-diet mice (Fig. 5H). Capsaicin treatment, either alone or combined with rimegepant, reduced the accumulation of myCAFs and type I collagen deposition, and increased immune cell infiltration (Fig. 5I-5K). In the 4T1 orthotopic tumor model, knockdown of Ramp1 in CAFs counteracted the accelerated tumor growth induced by capsaicin (Fig. 5L), reduced type I collagen deposition, and increased CD8⁺ T cell infiltration. In immune-competent C57BL/6J mice, Ramp1 knockdown reduced AT3 tumor burden and improved TME (Fig. 5M), but no such effect was observed in immune-deficient B-NDG mice, highlighting the critical role of RAMP1⁺ CAFs in responding to CGRP to regulate TME.
Fig. 5. Sensory neuron-derived CGRP binds to RAMP1 on CAFs to promote collagen secretion
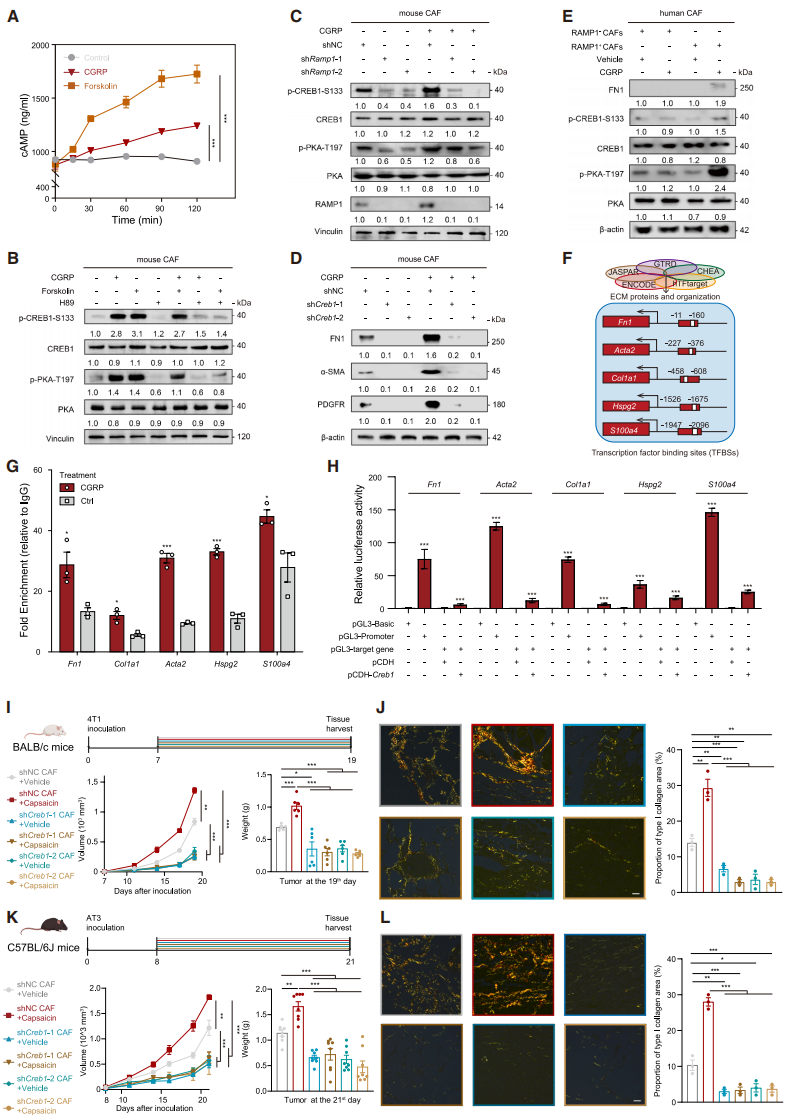
CGRP-RAMP1 Axis Enhances CAF Collagen Biosynthesis via cAMP/PKA/CREB1 Pathway
It is known that RAMP1 activation triggers adenylyl cyclase (AC) to produce cAMP, which then promotes the phosphorylation of cAMP response element-binding protein 1 (CREB1) via PKA. After CGRP treatment, cAMP production in CAFs increased, an effect similar to PKA agonist forskolin (Fig. 6A); PKA inhibitor H89 blocked both CGRP- and forskolin-induced p-PKA and p-CREB1 (Fig. 6B). In shRamp1 CAFs, p-PKA and p-CREB1 levels were reduced, and CGRP treatment could not restore these levels (Fig. 6C). Knockdown of CREB1 suppressed collagen synthesis and reduced the expression of myCAF markers (Fig. 6D). After CGRP treatment, RAMP1⁺ CAFs derived from human TNBC tissues exhibited increased levels of p-PKA, p-CREB1, and FN1 (Fig. 6E), confirming that CGRP activates CAFs via the cAMP/PKA/CREB1 pathway.
To explore the mechanism by which CREB1 regulates collagen synthesis, they identified potential binding sites (TFBSs) for CREB1 in multiple ECM-related gene promoters (Fig. 6F). ChIP-qPCR showed that CREB1 significantly enriched these TFBSs, with a more pronounced effect after CGRP treatment (Fig. 6G). In HEK293T cells, overexpression of CREB1 activated the luciferase activity of ECM-related genes, confirming that CREB1 directly regulates the transcription of ECM-related genes (Fig. 6H). In vivo experiments showed that 4T1 tumors carrying shCreb1 CAFs alleviated the tumor growth acceleration induced by sensory neuron activation, reversing fibrosis and the immune-exclusion TME (Fig. 6I, 6J); similar results were observed in models using AT3 cells and shCreb1 CAFs (Fig. 6K, 6L). In conclusion, CGRP-mediated CREB1 activation upregulates myofibroblast levels in CAFs and promotes tumor progression through dense collagen matrix formation.
Fig. 6. CGRP-RAMP1 axis enhances collagen biosynthesis in CAFs through the cAMP/PKA/CREB1 pathway
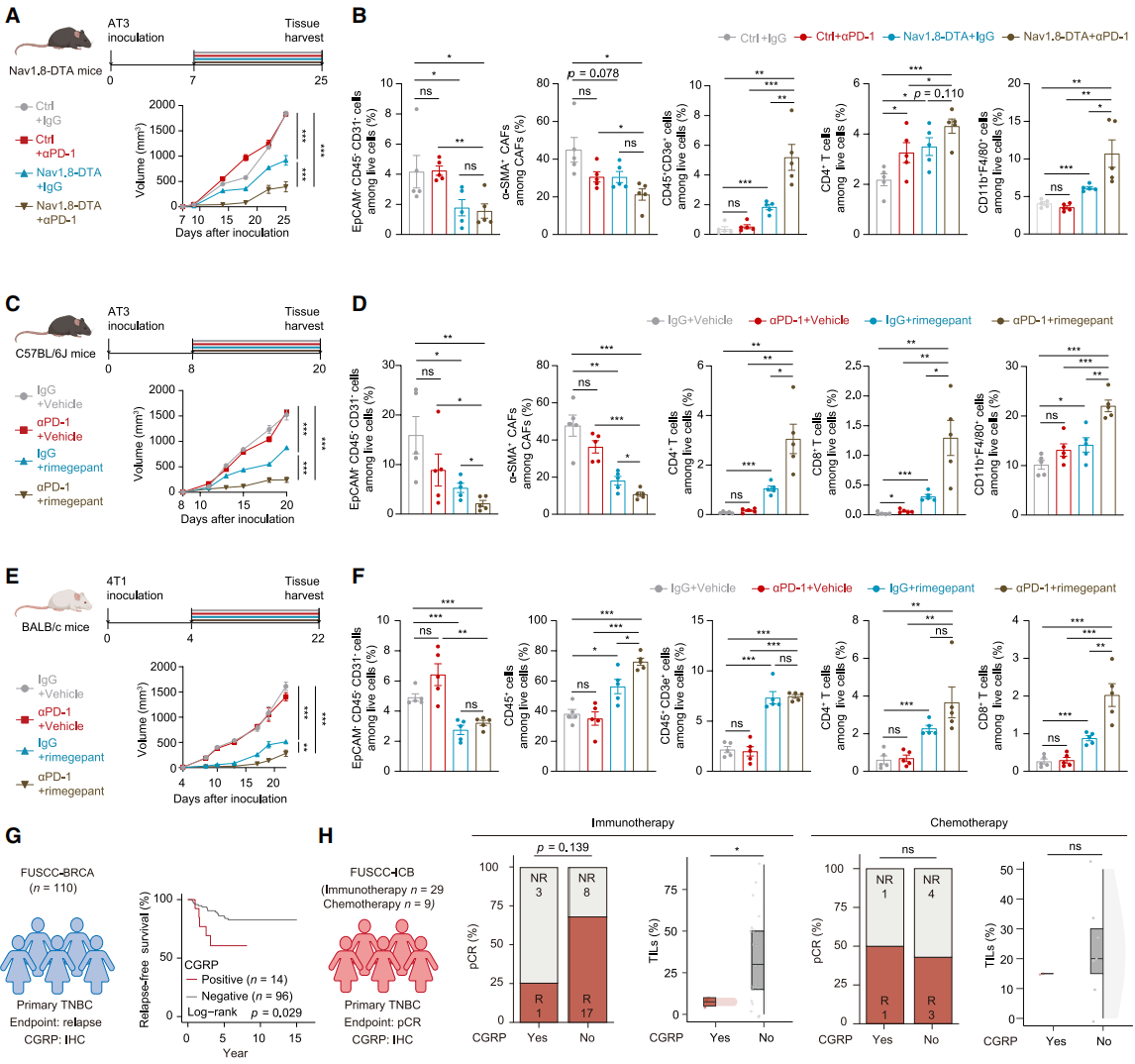
Based on the discovery that sensory neuron depletion improves TME, the researchers tested its synergistic effect with immunotherapy. An AT3 xenograft model was established in Nav1.8-DTA mice and their littermate controls, followed by PD-1 antibody treatment. Results showed that the sensory neuron ablation combined with anti-PD-1 group had the slowest tumor growth (Fig. 7A). In Nav1.8-DTA mice, the proportion of CAFs was lower, and immune cell infiltration increased after combining with anti-PD-1 (Fig. 7B). In C57BL/6J mice, rimegepant combined with anti-PD-1 treatment inhibited tumor growth, reduced myCAF differentiation and type I collagen deposition, and increased tumor-infiltrating immune cytotoxic cells (Fig. 7C, 7D), with similar results observed in BALB/c mice (Fig. 7E, 7F). Immunohistochemistry (IHC) staining confirmed that combination treatment increased CD8⁺ T cell infiltration. They then investigated the clinical significance of CGRP: using the FUSCC-BRCA cohort with 110 TNBC tissue samples, IHC staining confirmed that CGRP is a reliable biomarker for poor prognosis in TNBC patients (Fig. 7G). In the FUSCC-ICB neoadjuvant immunotherapy cohort, CGRP-positive patients showed a trend toward worse immunotherapy response, with fewer TILs infiltration, while chemotherapy efficacy was not affected by CGRP status (Fig. 7H). In conclusion, combining CGRP inhibitors with anti-PD-1 may be an ideal strategy for TNBC immunotherapy, and CGRP is expected to become a predictive biomarker for patient prognosis and immunotherapy response.
Fig. 7. Synergistic effect of targeting sensory neurons with ICB in TNBC
Conclusion
This study reveals that sensory neurons are the predominant neuronal type in the tumor microenvironment of triple-negative breast cancer. Tumor-secreted nerve growth factor activates sensory neurons to release CGRP, which binds to the RAMP1 receptor on cancer-associated fibroblasts. Through the cAMP/PKA/CREB1 signaling pathway, CGRP promotes collagen deposition, leading to the formation of a dense extracellular matrix that causes immune exclusion. Targeting sensory neurons can remodel the TME, synergizing with anti-PD-1 therapy to enhance efficacy, offering a new strategy for TNBC immunotherapy. Additionally, CGRP may serve as a biomarker for TNBC prognosis and immune therapy response.
The viral tools used in this study are available from Brain Case, which also offers various customized services. For more information, please contact