Client Articles | Neuron | Can’t sleep well under stress? Air Force Medical University team (Hailong Dong, Guangchao Zhao) reveals VTA–mPFC pathway drives sleep architecture disruption under chronic stress
Release time:2026-07-01 16:18:58
Chronic stress is widely known to induce sleep disturbances, characterized by sleep fragmentation and impaired sleep spindles—core comorbid features of psychiatric disorders such as depression. The ventral tegmental area (VTA) dopamine system is a key stress-responsive brain region, yet the causal mechanisms by which the VTA–mPFC dopaminergic circuit regulates sleep spindles and sleep stability remain unclear.
On May 20, 2026, a collaborative team led by Hailong Dong and Guangchao Zhao from Xijing Hospital, Air Force Medical University, published a study in Neuron titled “Hyperexcitable VTA-mPFC dopaminergic circuit disrupts sleep spindle and mediates sleep fragmentation after chronic social defeat stress.”
The study systematically elucidates how chronic social defeat stress (CSDS) disrupts the VTA→mPFC dopaminergic circuit, leading to impaired sleep spindles and fragmented NREM sleep. The authors provide multi-level validation of this mechanism, spanning behavioral phenotypes, electrophysiology, circuit projections, neurotransmitter release, and ion channel targets.
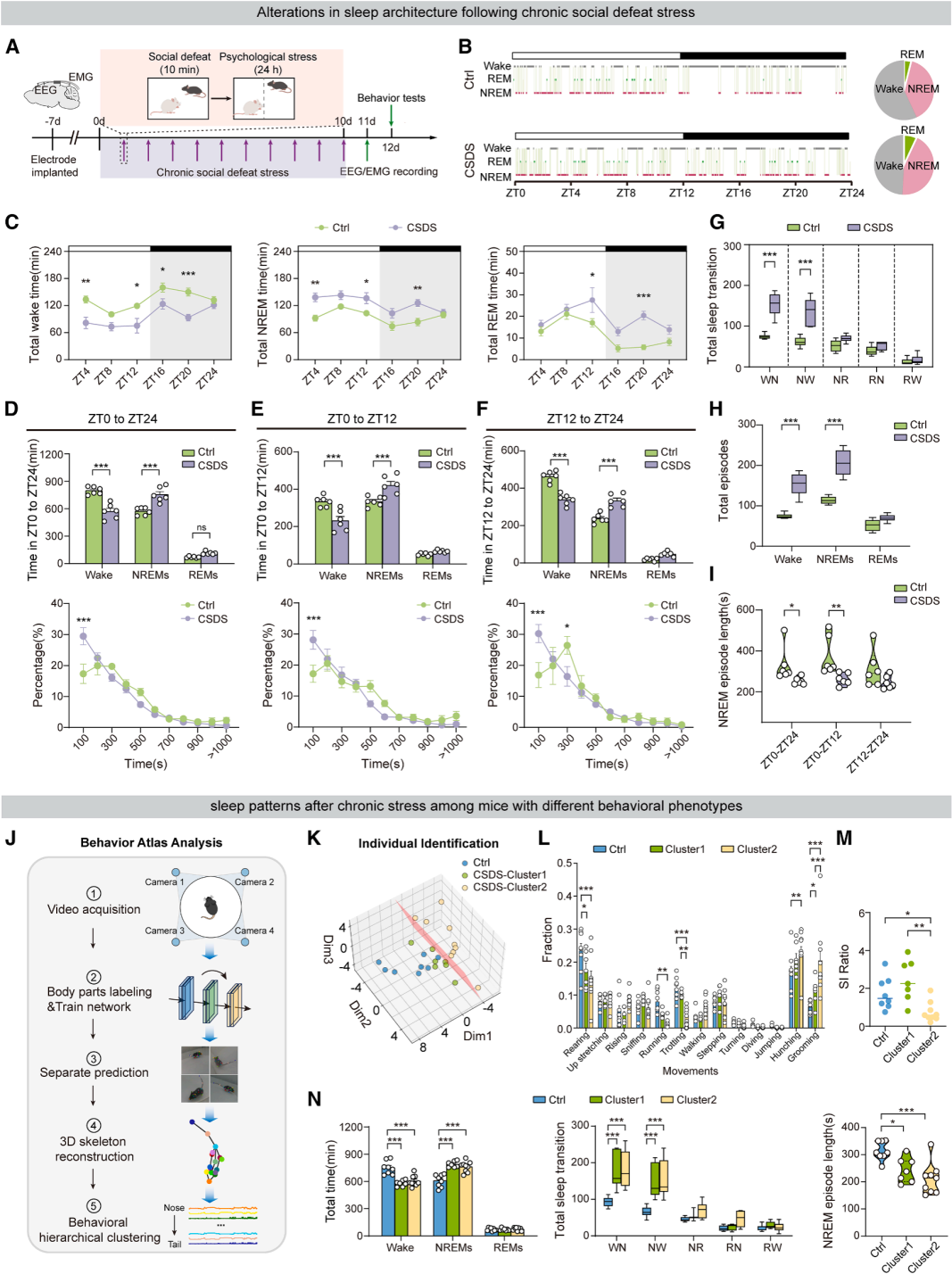
Part 01. CSDS increases but fragments non-rapid eye movement sleep (NREMS)
In this study, a chronic social defeat stress (CSDS) mouse model was used in combination with wireless telemetry EEG/EMG recordings to investigate how chronic stress affects sleep architecture. C57BL/6J mice were subjected to 10 minutes of daily aggression from CD-1 aggressive male mice, followed by 24 hours of continuous sensory contact with the aggressor (Fig. 1A). After the stress paradigm, all mice underwent 24-hour continuous sleep monitoring.
The results showed that CSDS mice exhibited increased total NREM sleep duration and reduced wakefulness, but significantly impaired sleep continuity. Meanwhile, the frequency of wake-to-NREM transitions and NREM episodes increased markedly, along with a higher number of sleep state transitions. The average duration of individual NREM bouts was reduced, indicating clear NREM sleep fragmentation (Fig. 1B–1I).
To capture individual behavioral heterogeneity, CSDS mice were further classified into two phenotypic subgroups (CSDS-1 and CSDS-2) (Fig. 1J–1K). All stressed mice showed reduced exploratory behavior and increased stereotyped behaviors, with significant behavioral variability between the two groups (Fig. 1L–1M). Importantly, regardless of behavioral phenotype, stress susceptibility, or resilience, all CSDS mice exhibited a consistent sleep disturbance profile, characterized by increased NREM duration and enhanced sleep fragmentation. These findings indicate that stress-induced alterations in sleep architecture represent a stable and generalizable physiological phenotype, independent of individual differences in emotional or social behavior.
Figure 1. Alterations in sleep architecture following CSDS
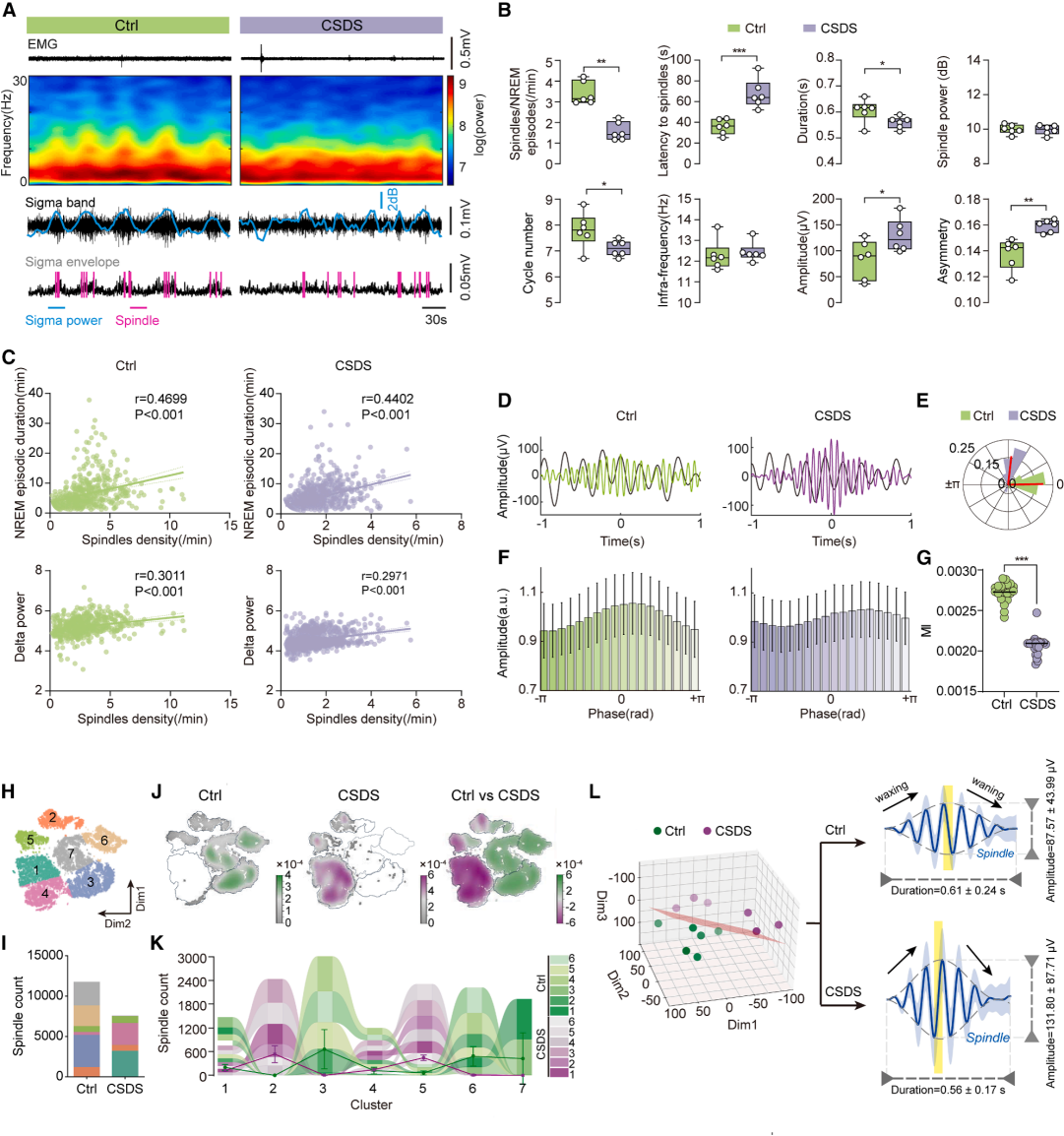
Part 02. CSDS disrupts spectral power and sleep spindle microstructure during NREM sleep
Spectral analysis revealed reduced sleep depth and impaired sleep quality in CSDS mice. Sleep spindles, which are predominantly observed during N2-stage sleep, were significantly disrupted following stress exposure. Stressed mice showed a marked reduction in spindle density, accompanied by prolonged latency and shortened duration, indicating that chronic stress impairs both the quantity and microstructure of sleep spindles (Fig. 2A–2B).
Spindle density was positively correlated with NREMS stability and delta (δ) wave slow-wave activity, and this relationship remained intact after CSDS manipulation (Fig. 2C). Delta–spindle coupling is critical for sleep homeostasis and memory consolidation. CSDS shifted the phase relationship between spindle peaks and delta waves, reduced the modulation index (MI), and significantly weakened their temporal coordination (Fig. 2D–2G).
Further analysis of stress effects on spindle subtypes identified seven distinct spindle categories. The distribution of these subtypes differed markedly between control and CSDS groups, enabling accurate discrimination of stressed individuals based on spindle characteristics. Quantitative analyses further showed that CSDS mice exhibited increased spindle amplitude, reduced duration, and enhanced asymmetry, representing a distinct abnormal profile (Fig. 2H–2L). Notably, these spindle impairments were highly consistent across both stress-susceptible and stress-resilient mice, indicating that disruption of spindle microstructure is a stable and intrinsic physiological marker of chronic stress, independent of individual behavioral phenotypes.
Figure 2. Characteristics of sleep spindles following CSDS
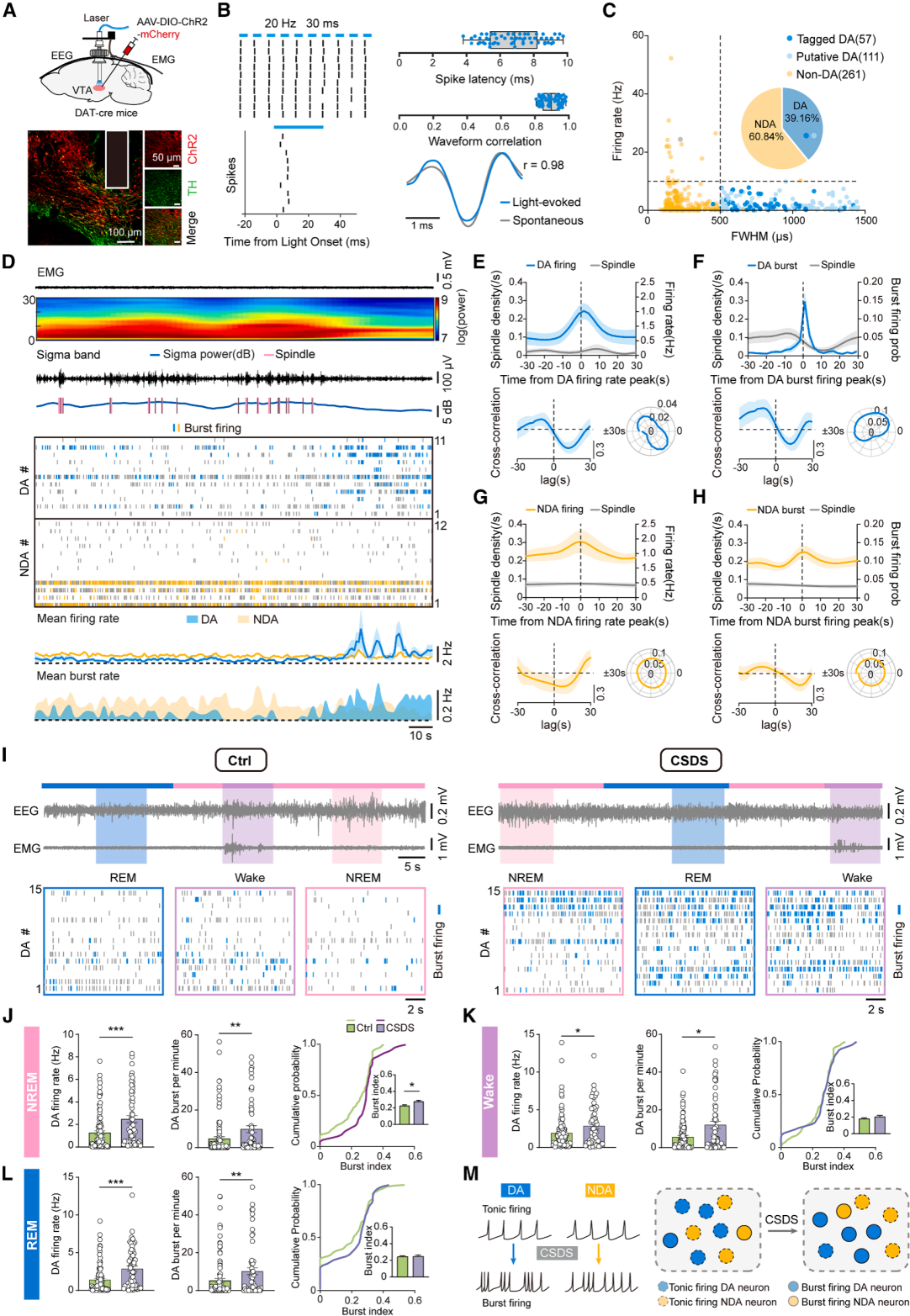
Part 03. Reduced sleep spindles are associated with abnormal firing of VTA dopaminergic neurons after CSDS
To investigate the underlying mechanism, in vivo recordings of ventral tegmental area dopamine (VTA^DA) neurons were performed using optetrode electrophysiology combined with optogenetic identification (Fig. 3A–3C). Temporal analysis revealed that high-frequency firing of VTA^DA neurons—particularly burst firing—significantly suppressed spindle generation and disrupted spindle continuity. Both firing rate and burst probability were negatively correlated with spindle density.
In contrast, activity of non-dopaminergic VTA neurons showed no significant correlation with spindle dynamics, indicating that spindle regulation specifically depends on VTA^DA neuronal activity (Fig. 3D–3H). After CSDS, VTA^DA neurons exhibited increased firing and bursting across all sleep stages, while burst activity increased specifically during NREM sleep (Fig. 3I–3L). Non-dopaminergic neurons also showed increased firing, but without changes in NREM burst index.
Taken together, CSDS selectively enhances burst firing of VTA^DA neurons during NREM sleep, which is a key driver of spindle deficits and sleep disruption.
Figure 3. CSDS enhances firing rate and burst activity of VTA^DA neurons, leading to abnormal sleep spindles
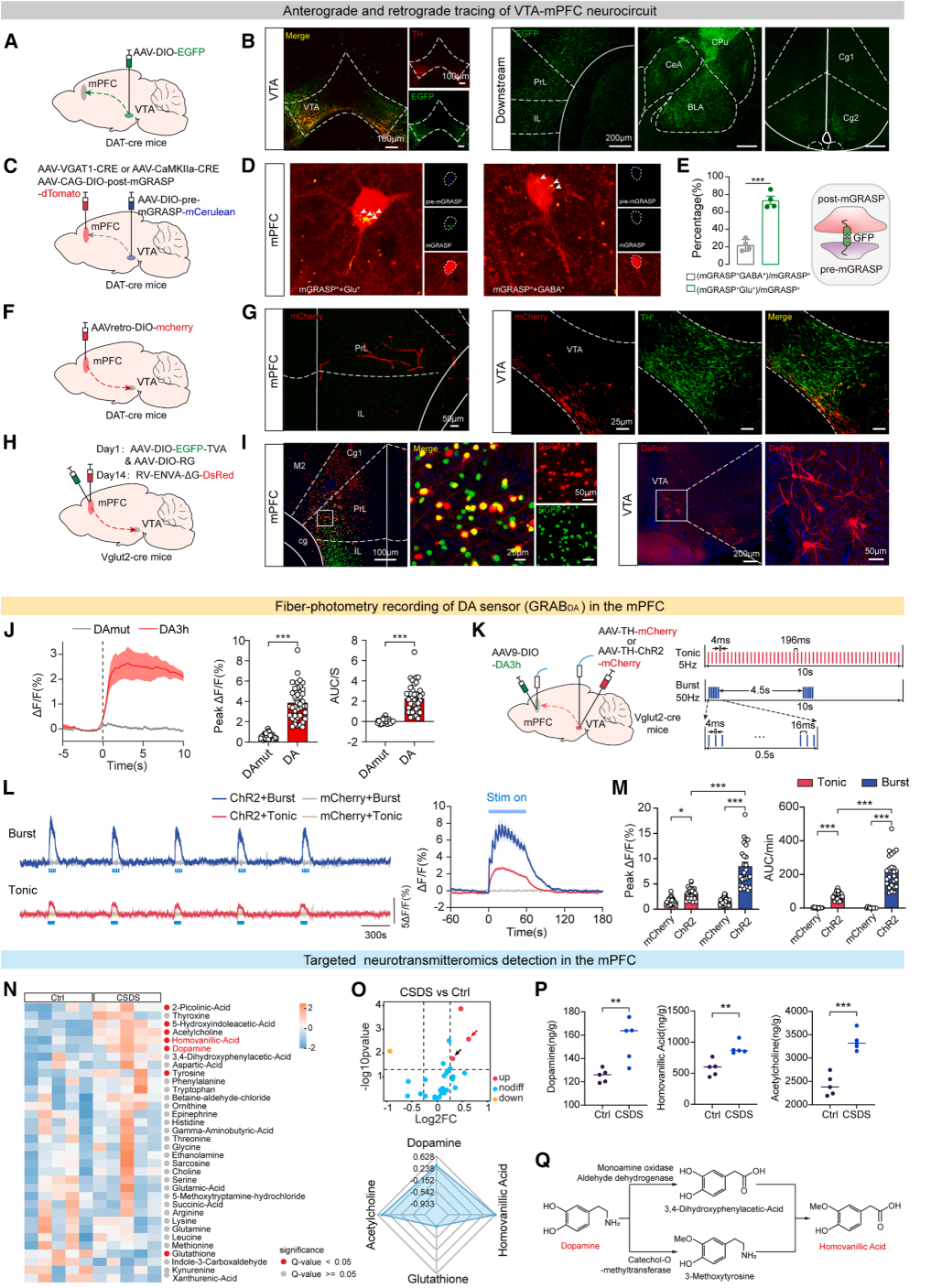
Part 04. CSDS-induced VTA dopamine burst firing leads to excessive dopamine release in the medial prefrontal cortex (mPFC)
Viral tracing experiments showed that VTA^DA fibers widely project to multiple brain regions, including the mPFC, amygdala, and striatum (Fig. 4A–4B). The study then focused on the core VTA–mPFC circuit. mGRASP synaptic labeling, retrograde viral tracing, and rabies virus-based monosynaptic tracing collectively demonstrated that VTA^DA neurons preferentially form synaptic connections with glutamatergic excitatory neurons in the mPFC (Fig. 4C–4I), defining the anatomical basis of a VTA→mPFC glutamatergic-associated dopaminergic circuit.
Calcium imaging and dopamine sensors further showed that stress significantly increased VTA neuronal activity, leading to robust elevation of dopamine transients in mPFC glutamatergic neurons, whereas GABAergic neurons exhibited only a modest increase, indicating strong cell-type specificity (Fig. 4J).
Optogenetic stimulation at matched intensities demonstrated that both tonic (5 Hz) and burst-like (50 Hz) firing patterns promoted dopamine release in the mPFC, with VTA^DA burst firing inducing stronger and more sustained dopamine elevation (Fig. 4K–4M).
Mass spectrometry confirmed that CSDS significantly increased levels of dopamine, its metabolite HVA, and acetylcholine in the mPFC (Fig. 4N–4Q). These findings demonstrate that chronic stress induces VTA^DA burst firing, resulting in excessive dopamine accumulation in the mPFC, which represents a central mechanism underlying stress-induced sleep disruption.
Figure 4. Pattern-specific optogenetic control of VTA^DA neurons and dopamine release in the mPFC after CSDS
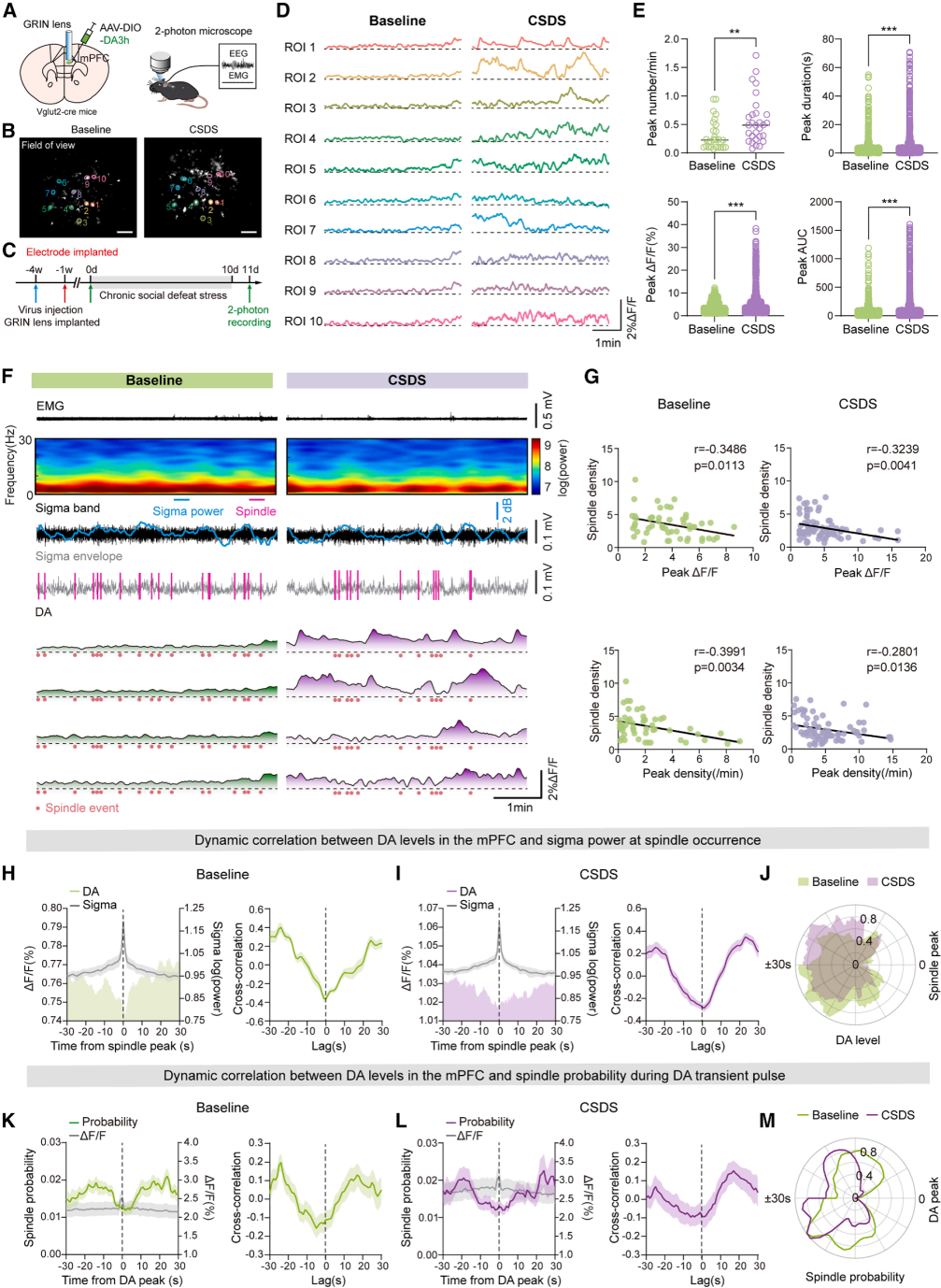
Part 05. Increased dopamine in the mPFC disrupts sleep spindle continuity
In vivo two-photon imaging combined with EEG recordings was used to monitor dopamine dynamics in the medial prefrontal cortex (mPFC) (Fig. 5A–5C). Under physiological conditions, dopamine levels fluctuate in a regular pattern during wake–NREM transitions. After CSDS, dopamine levels were abnormally elevated during both wakefulness and NREM sleep.
Stress increased the frequency and amplitude of dopamine transients during NREM sleep, with dopamine peaks preferentially occurring during spindle-free intervals. Dopamine dynamics and spindle-related measures showed a significant negative correlation (Fig. 5D–5G). Normally, sleep spindles occur at peaks of sigma power and low dopamine states, whereas transient dopamine elevations suppress spindle occurrence (Fig. 5H–5M). These findings suggest that excessive dopamine in the mPFC disrupts the continuous generation of sleep spindles.
Figure 5. Dopamine pulses interrupt spindle continuity after CSDS
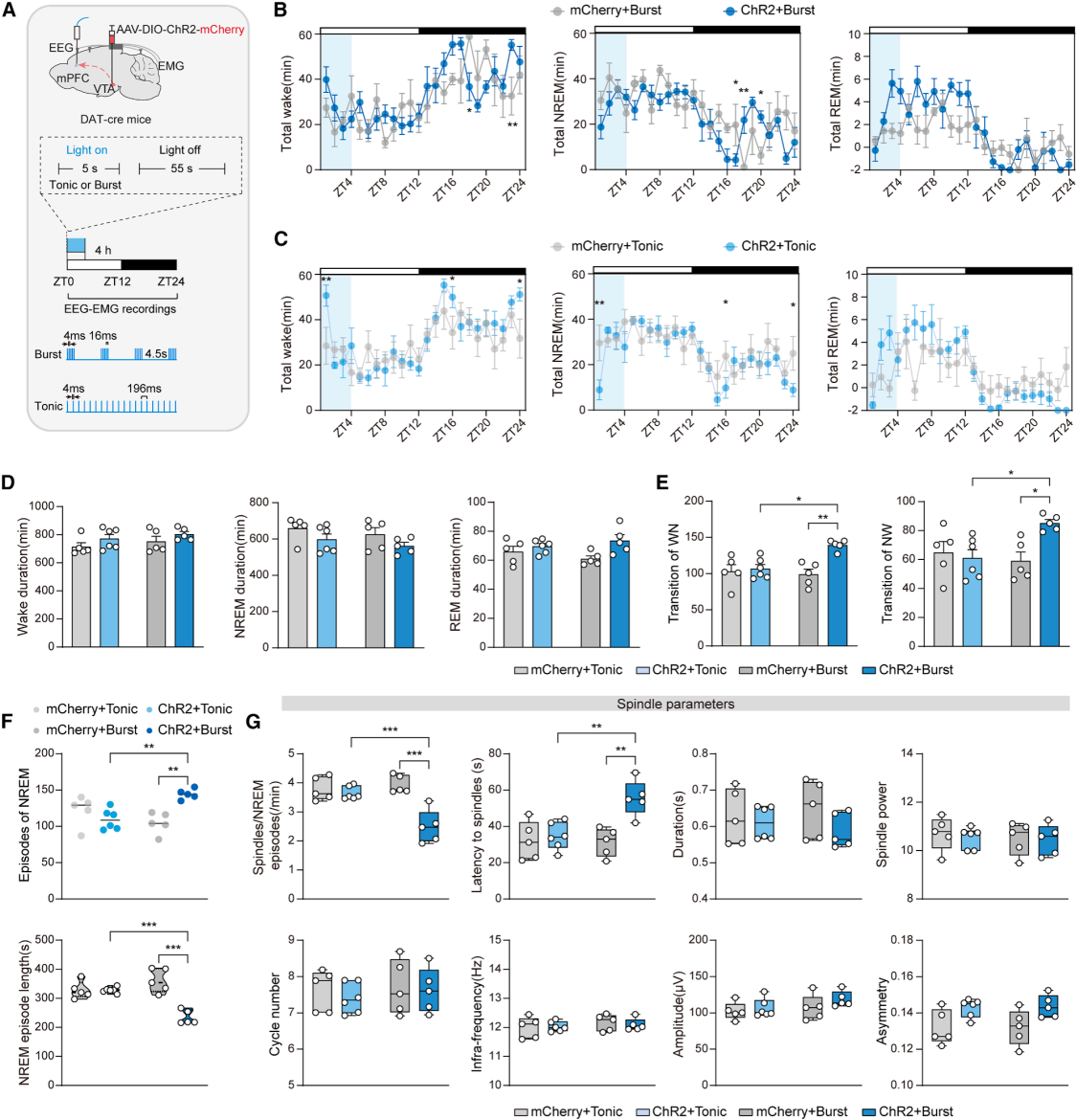
Part 06. Distinct firing patterns in the VTA–mPFC circuit induce sleep abnormalities
Using region-specific optogenetic stimulation of VTA→mPFC terminals (Fig. 6A), both tonic (5 Hz) and burst (50 Hz) stimulation paradigms were applied. Neither stimulation condition altered the overall proportion of total daily sleep (Fig. 6B–6D). However, only burst-pattern activation induced sleep fragmentation and impaired spindle development (Fig. 6E–6G), demonstrating that sleep regulation depends on firing pattern specificity within the circuit.
Figure 6. Burst activation of the VTA^DA–mPFC pathway induces sleep abnormalities
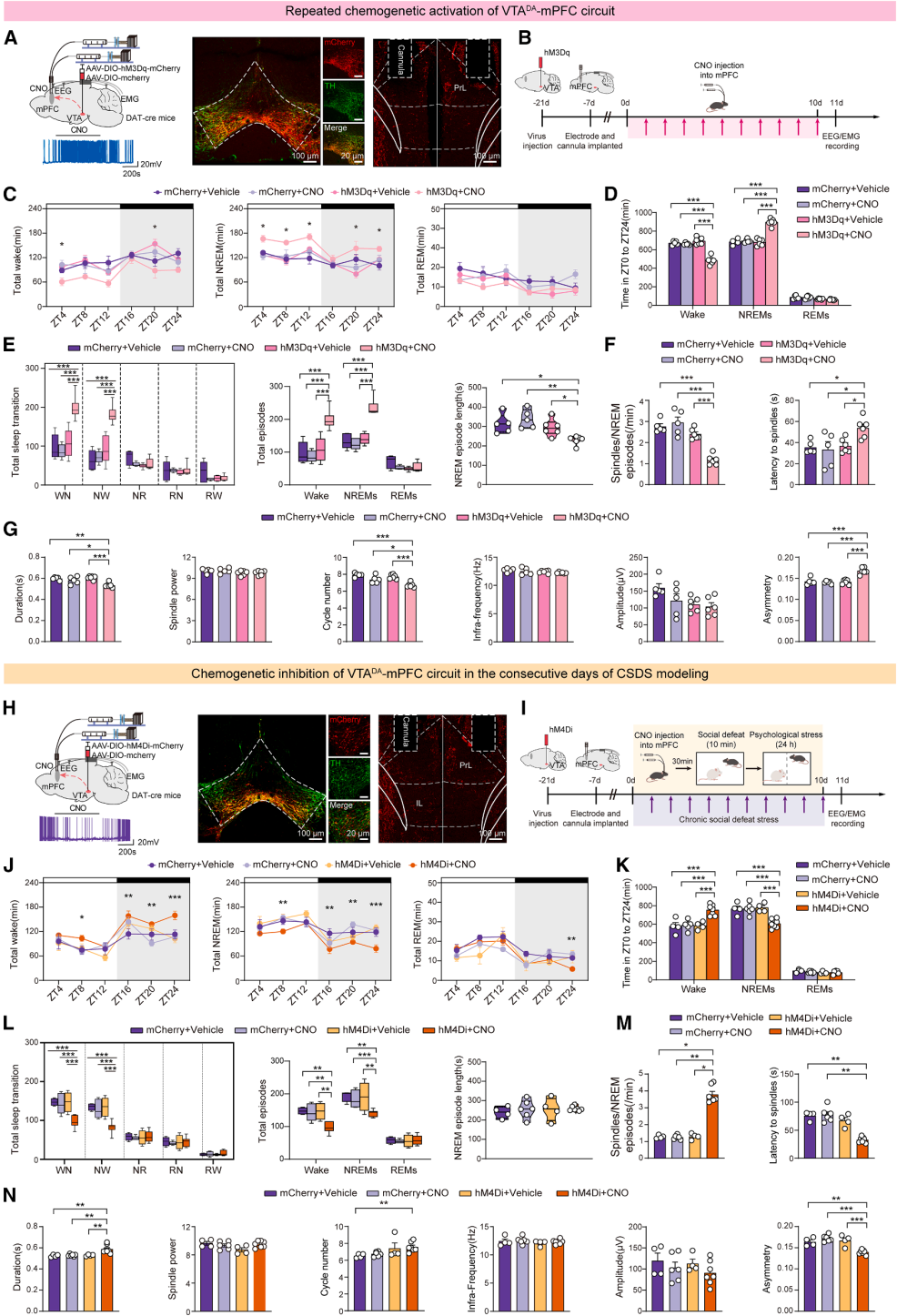
Part 07. Overactivation of the VTA–DA–mPFC circuit mediates stress-induced sleep architecture and spindle abnormalities
The study used the hM3Dq chemogenetic system to specifically activate VTA^DA neurons in mice (Fig. 7A–7B), which significantly increased dopamine levels in the mPFC. Sustained activation of this circuit was sufficient to recapitulate the characteristic sleep impairments observed in CSDS, including reduced wakefulness, increased NREM duration, frequent sleep state transitions, and fragmented NREM episodes (Fig. 7C–7E).
At the same time, spindle abnormalities were induced, including reduced spindle density, shortened duration, prolonged latency, and increased structural asymmetry (Fig. 7F–7G). These results demonstrate that overactivation of the VTA^DA–mPFC circuit is sufficient to independently induce stress-like sleep disturbances and spindle deficits.
The study further applied the hM4Di chemogenetic inhibition system to suppress VTA^DA–mPFC circuit activity during CSDS modeling (Fig. 7H–7I). Circuit inhibition significantly alleviated stress-induced sleep disruption and spindle impairments, effectively improving sleep fragmentation and restoring multiple spindle-related parameters (Fig. 7J–7N). However, it did not rescue deficits in the average duration of individual NREM bouts.
Taken together, overactivation of the VTA^DA–mPFC circuit is both necessary and sufficient for mediating stress-induced sleep architecture disruption and spindle dysfunction.
Figure 7. The VTA^DA–mPFC projection regulates stress-induced sleep architecture and spindle abnormalities
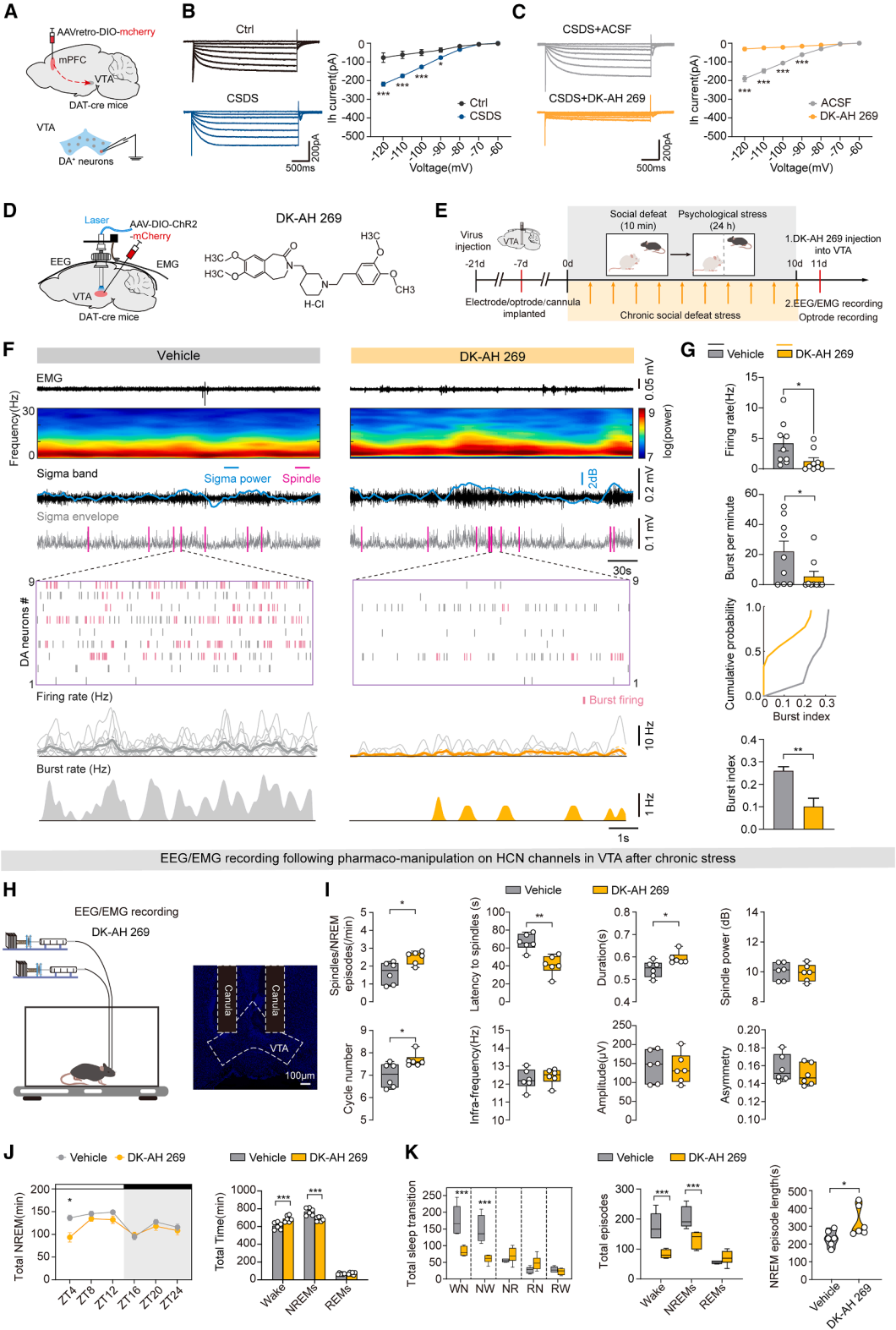
Part 08. HCN channel inhibition suppresses VTA dopamine burst firing and alleviates CSDS-induced sleep disturbances
Hyperpolarization-activated cyclic nucleotide-gated (HCN) channel–mediated Ih currents regulate burst firing in these neurons. The study specifically labeled and targeted a subpopulation of VTA dopamine neurons projecting to the mPFC.
Patch-clamp recordings showed that CSDS significantly enhanced Ih currents in these neurons (Fig. 8A–8B). The HCN channel inhibitor DK-AH 269 effectively blocked Ih currents, confirming that HCN channels mediate stress-related neuronal abnormalities (Fig. 8C).
In vivo electrophysiological recordings further demonstrated that DK-AH 269 significantly reversed CSDS-induced increases in firing rate and burst activity of VTA dopamine neurons (Fig. 8D–8G). Behavioral and sleep monitoring after pharmacological intervention showed that DK-AH 269 markedly restored multiple spindle parameters, improved overall sleep architecture, and enhanced NREM sleep stability (Fig. 8H–8K).
In summary, CSDS activates the HCN–Ih pathway in VTA dopamine neurons, driving excessive burst firing that leads to spindle disruption and sleep fragmentation. Pharmacological blockade of HCN channels corrects abnormal neuronal firing and effectively alleviates chronic stress–induced sleep disturbances.
Figure 8. DK-AH 269 suppresses burst firing of VTA^DA neurons and improves chronic stress–induced sleep impairments
Part 09. Summary
Chronic stress enhances burst firing in the VTA→mPFC dopaminergic circuit, leading to excessive dopamine in the prefrontal cortex, disruption of sleep spindles, and ultimately fragmented NREM sleep. Inhibition of this circuit or blockade of HCN channels can reverse stress-induced sleep disturbances.
The tools used in this article are all available from Brain Case Biotech: