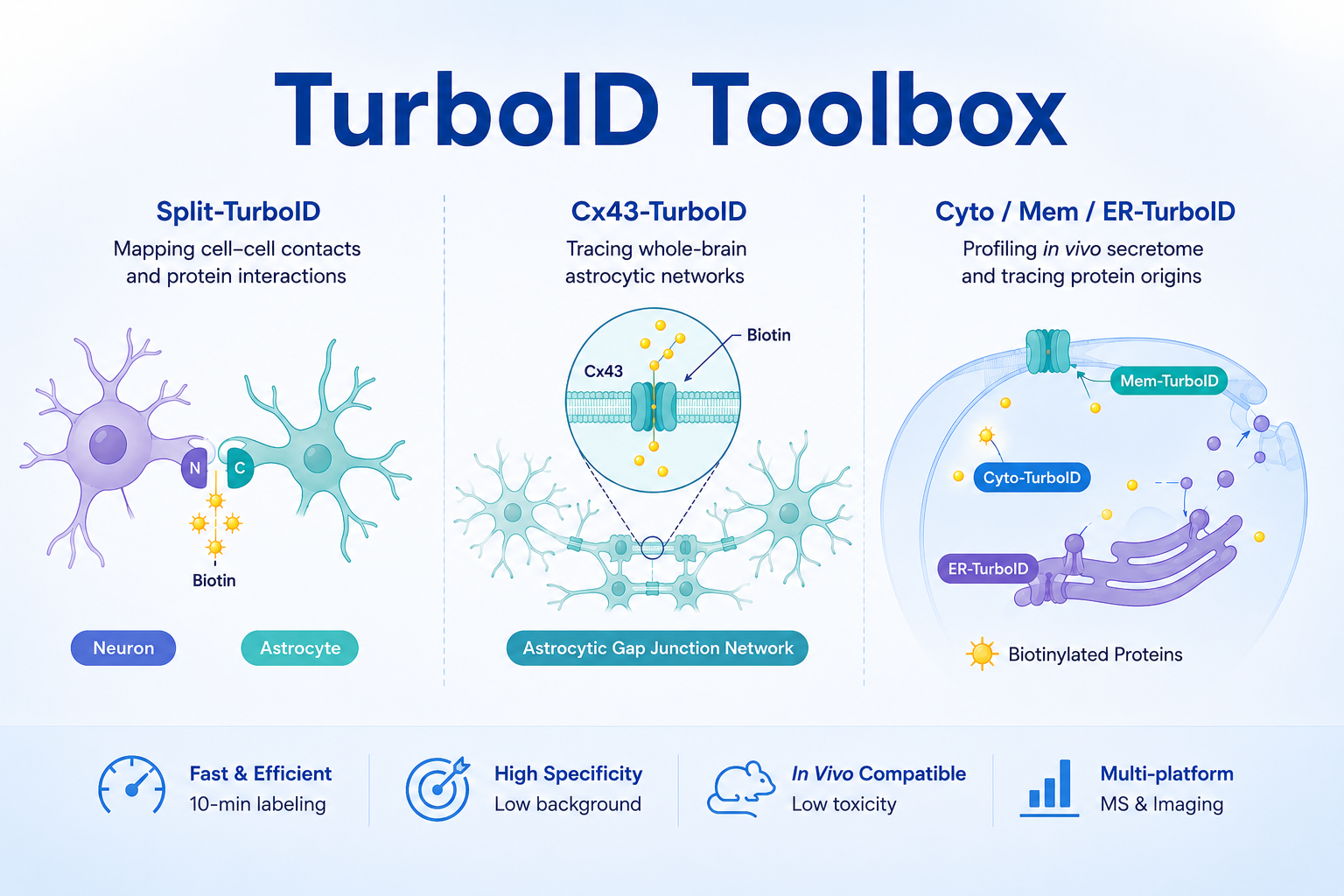
TurboID Toolbox | Three TurboID-Derived Tools for Comprehensive Analysis of Cell–Cell Interactions, Astrocytic Networks, and the In Vivo Secretome
Release time:2026-07-15 16:14:14
Proximity labeling (PL) relies on a target-fused promiscuous biotin ligase to covalently label proteins within a nanometer-scale radius upon the addition of exogenous biotin. Combined with streptavidin enrichment and mass spectrometry, PL enables the analysis of protein–protein interactions and organelle proteomes.
There are two major classes of proximity labeling tools currently in use:
APEX:Labeling can be completed within 1 minute, but it requires toxic H₂O₂, which limits its application in living organisms.
BioID (BirA-R118G):Requires only the addition of exogenous biotin and exhibits low cytotoxicity. However, labeling requires 18–24 hours, making it incapable of capturing transient and dynamic protein interactions. Its derivative variants, BioID2 and BASU, exhibit catalytic activities comparable to that of BioID but still require prolonged labeling times, limiting their ability to achieve effective labeling in model organisms such as Caenorhabditis elegans and Drosophila melanogaster. In 2018, the laboratory of Alice Y. Ting published a study in Nature Biotechnology, in which the Escherichia coli biotin ligase BirA was engineered through yeast display-directed evolution to develop two novel proximity labeling enzymes, TurboID and miniTurbo. These enzymes substantially overcome the major limitation of conventional BioID—its prolonged labeling time—and enable rapid and efficient proximity protein labeling in living cells and living organisms.
I. Enzyme Origin and Basic Properties
TurboID is a highly active mutant of the wild-type Escherichia coli biotin ligase BirA, generated through multiple rounds of yeast display-directed evolution (35 kDa, containing 15 amino acid substitutions). Its derivative, miniTurbo (28 kDa, generated by deleting the N-terminal amino acids 1–63 and introducing 13 amino acid substitutions), is a truncated version of TurboID. Both enzymes are promiscuous biotin ligases. Compared with the first-generation BioID (BirA-R118G), their catalytic activity is increased by tens of fold, enabling efficient labeling after only 10 minutes of exogenous biotin supplementation.
II. Core Biochemical Catalytic Mechanism
🌟Step 1: Generation of the Activated Intermediate
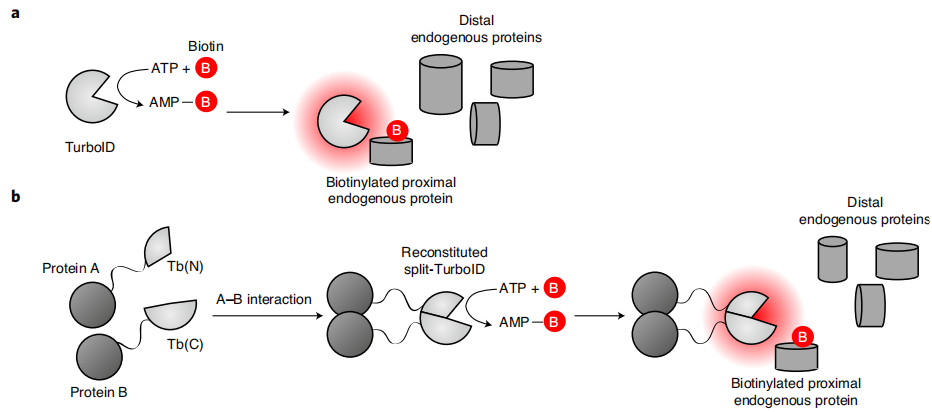
TurboID binds free biotin and intracellular ATP as substrates and catalyzes the formation of the high-energy intermediate biotin-5′-AMP (biotinyl-5′-adenylate). This activated molecule does not remain covalently bound to the enzyme, but instead diffuses freely out of the TurboID catalytic pocket, with an effective diffusion radius of approximately 10 nm.
🌟Step 2: Covalent Biotinylation of Neighboring Proteins
The released biotin-5′-AMP is highly reactive and spontaneously forms covalent bonds with the amino groups of exposed lysine (Lys) side chains on neighboring proteins, thereby stably biotinylating neighboring proteins. Only proteins located within approximately 10 nm of TurboID are labeled, whereas proteins located farther away undergo little or no modification, thereby achieving proximity-dependent labeling. This process does not require direct physical interaction between proteins; spatial proximity alone is sufficient for labeling. Characteristics: Biotin labeling is a covalent modification, allowing the biotin signal to remain stable after cell lysis and protein denaturation, which is essential for subsequent streptavidin enrichment.Only the addition of exogenous biotin is required, without the need for toxic reagents such as H₂O₂ (in contrast to APEX), resulting in very low toxicity in living cells and living animals. The native labeling radius of approximately 10 nm represents the characteristic spatial resolution of TurboID. Modification of the linker can fine-tune the labeling radius; excessively long linker peptides increase the labeling range and lead to a higher rate of false-positive labeling.
Figure 1 | Proximity-dependent biotinylation reactions catalyzed by TurboID and split-TurboID
III. Principles of Three Common TurboID-Derived Systems
🌟1. Split-TurboID (A Tool for Studying Astrocyte Synapses)
TurboID is split into two catalytically inactive fragments, an N-terminal fragment (N-Turbo) and a C-terminal fragment (C-Turbo). These two fragments are separately driven by cell type-specific promoters in different cell types (neurons or astrocytes) and fused to a glycosylphosphatidylinositol (GPI) anchor, thereby directing them to the outer surface of the plasma membrane and avoiding interference from intracellular signals. Only when the two cell types establish membrane contact and the two fragments are brought into close spatial proximity do they spontaneously reconstitute into an enzymatically active TurboID. The activated intermediate is therefore generated exclusively at the cell–cell contact interface, resulting in the labeling of proteins across the intercellular cleft. This system is specifically designed for the identification of proteomes at cell–cell contact sites, such as the astrocyte–neuron synaptic cleft.
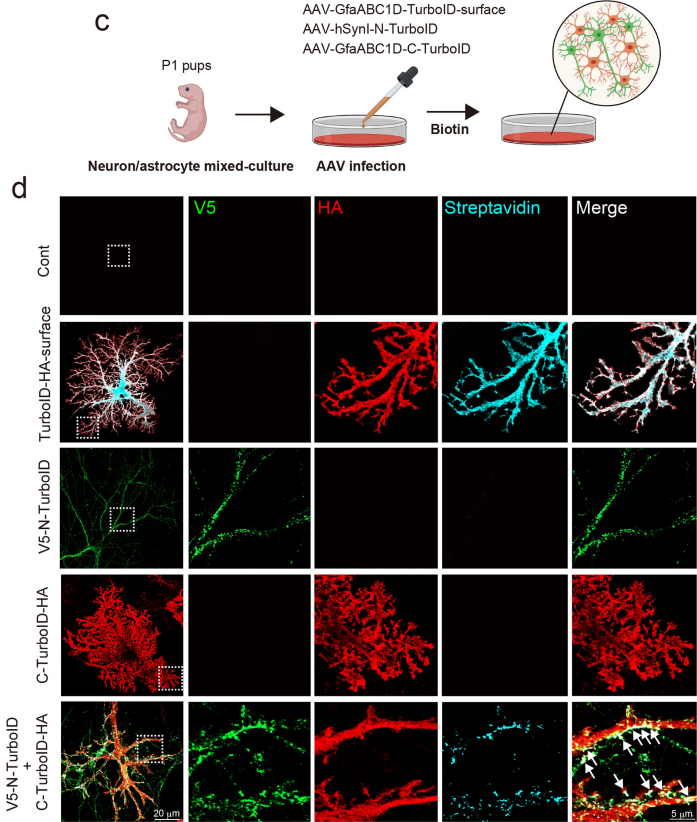
💎 (1) In Vitro Cell Validation
Neuron–astrocyte co-culture system: No biotin labeling was detected when either N-Turbo or C-Turbo was expressed alone. Specific biotin labeling was observed only when the two cell types were co-cultured and membrane contact was established. Fluorescence colocalization analysis showed that the labeling signals were highly colocalized with markers of excitatory synapses (VGLUT/PSD95) and inhibitory synapses (VGAT/gephyrin), demonstrating that the system specifically targets the tripartite synaptic cleft.
Figure 2 | Reconstituted activity of Split-TurboID in cultured neurons and astrocytes
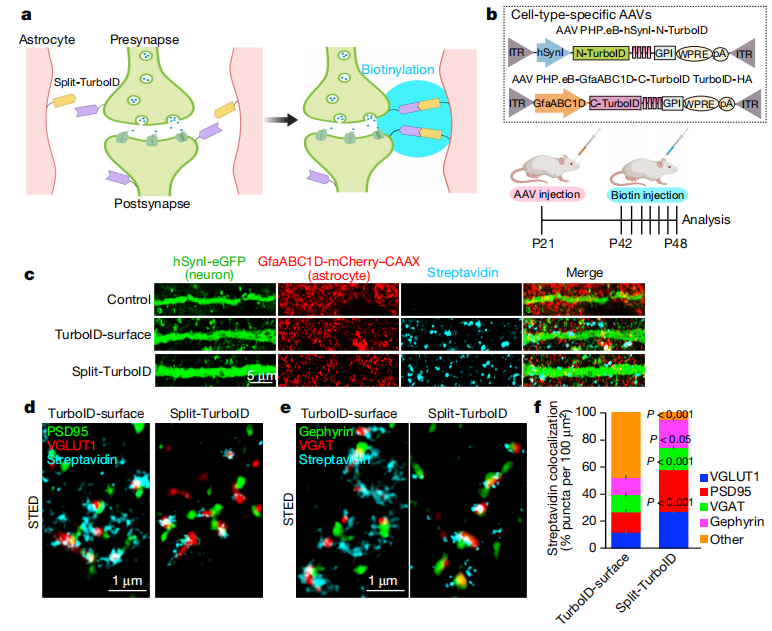
💎 (2) In Vivo Labeling and Proteomic Profiling in Mice
P21 mice received retro-orbital co-injection of AAV-PHP.eB-hSynI-V5-TurboID(N)-GPI and AAV-PHP.eB-GfaABC1D-TurboID(C)-HA-GPI. The mice were maintained for 3 weeks to allow sufficient viral expression. Beginning at P42, biotin was administered by subcutaneous injection for 7 consecutive days to initiate in vivo labeling. A full-length Turbo-surface construct (GPI-anchored across the entire astrocyte plasma membrane, enabling global membrane labeling) was included as the control. The results showed that Split-Turbo labeling was enriched in the perisynaptic region. Stimulated emission depletion (STED) super-resolution microscopy demonstrated that more than 90% of the labeling signals were located immediately adjacent to synapses, whereas approximately 50% of the Turbo-surface signals were tightly colocalized with synaptic markers.
Figure 3 | Identification of astrocyte–neuron synaptic cleft proteomes using in vivo Split-TurboID
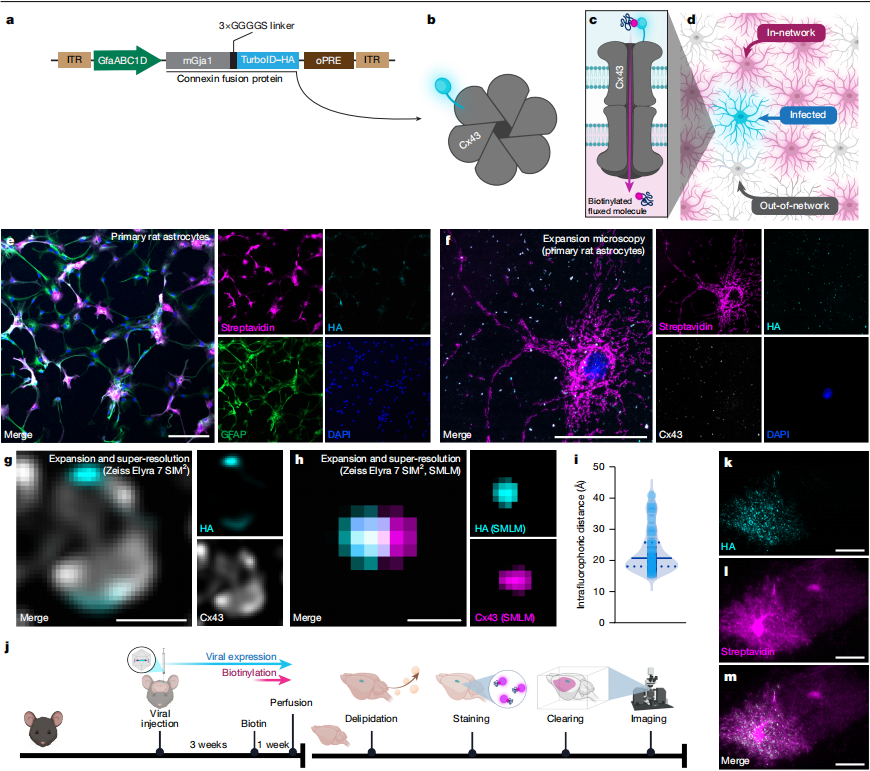
🌟2. Cx43-TurboID (A Whole-Brain Astrocyte Tracing Tool, Nature 2026)
Astrocytes form a syncytial network through Cx43/Cx30 gap junctions, which participate in brain metabolism, synaptic plasticity, and neural injury repair. Full-length TurboID is fused to the astrocyte-specific gap junction protein Cx43 via a flexible 3×GGGGS linker. The fusion protein is incorporated into the connexon of the gap junction (with TurboID fused to one of the six Cx subunits), thereby restricting TurboID to the lumen of the gap junction channel. As small molecules pass through the gap junction, they are biotinylated by TurboID within the channel lumen. The diffusion of these biotinylated small molecules is then used to trace the whole-brain astrocytic gap junction network.
✦ (1) In Vitro Validation in Primary Cells
Primary rat astrocytes were transduced with AAV expressing Cx43-TurboID. When only 10% of the cells expressed Cx43-TurboID, more than 80% of the cells were labeled by streptavidin. In astrocytes with Cx43/Cx30 double knockout, labeling was strictly restricted to the infected cells and failed to spread to neighboring cells, confirming the specificity of the system. Expansion microscopy combined with single-molecule imaging further demonstrated that TurboID was precisely localized within the lumen of the gap junction.
✦(2) Topological Analysis of the Whole-Brain Astrocytic Network in Mice
AAV was stereotaxically injected unilaterally into the motor cortex, prefrontal cortex, and hypothalamus of mice, respectively. After 3 weeks of viral expression, biotin was supplied in the drinking water for 7 days to initiate labeling. On Day 28, mice were transcardially perfused, and the brains were collected for delipidation, staining, and tissue clearing. Three-dimensional imaging was then performed using light-sheet microscopy, followed by quantitative analysis through registration to the Allen Mouse Brain Atlas. The results showed characteristic punctate HA signals corresponding to gap junctions on the plasma membrane of infected astrocytes. Streptavidin labeling was also detected in neighboring astrocytes that were not infected but belonged to the same astrocytic network. Although these cells did not express the fusion protein, they contained biotinylated small molecules that had diffused through gap junctions.
Figure 4 | Visualization of Astrocytic Gap Junction Communication Using an Astrocyte Network Tracer
🌟3. Cyto-TurboID / Mem-TurboID / ER-TurboID (Three Subcellular Localization Systems for In Vivo Secretome Profiling)
By incorporating different targeting signal peptides, TurboID is localized to distinct subcellular compartments, enabling precise discrimination of secretory pathways at their origin. The labeling ranges of the three systems do not overlap and are fully complementary.
💎 1. ER-TurboID (Endoplasmic Reticulum-Targeted): ER-TurboID is generated by fusing an N-terminal endoplasmic reticulum signal peptide with an ER retention sequence, thereby specifically anchoring TurboID within the lumen of the endoplasmic reticulum. It exclusively labels newly synthesized secretory proteins that enter the classical ER–Golgi secretory pathway and contain signal peptides, making it a dedicated tool for profiling the classical secretome.
💎 2. Mem-TurboID (Plasma Membrane-Targeted): Mem-TurboID is generated by fusing the transmembrane domain of PDGFRβ, thereby anchoring TurboID to the inner surface of the plasma membrane. It simultaneously labels two classes of secreted substrates: proteins secreted through the classical secretory pathway and extracellular domains of membrane receptors released by ectodomain shedding, thereby filling the gap in the detection of membrane-shed secreted molecules. 💎 3. Cyto-TurboID (Cytosolic): Cyto-TurboID contains no targeting signal peptide and is freely distributed throughout the cytosol. It does not enter the endoplasmic reticulum pathway and therefore serves as the only specific tool for profiling the unconventional secretory pathway.
✦ (1) In Vitro Cell Validation
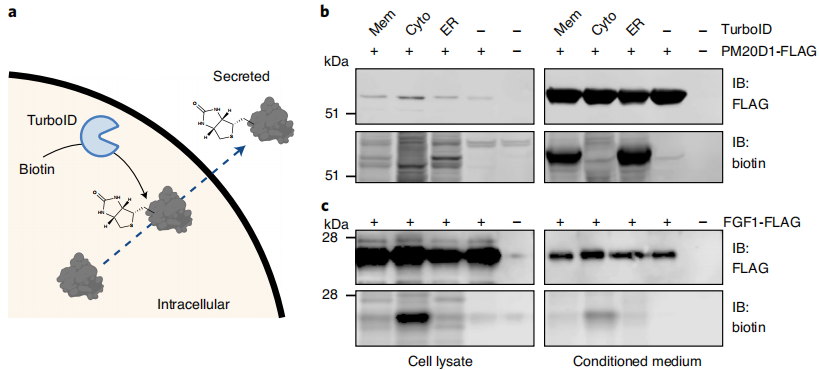
Validation was performed in HEK293T cells by co-transfecting each TurboID system with known secreted proteins to determine the labeling preference of each system. The classical secretory protein PM20D1 was efficiently labeled only by ER-TurboID and Mem-TurboID, whereas Cyto-TurboID produced almost no detectable signal. In contrast, the unconventional secretory protein FGF1 was enriched exclusively in the Cyto-TurboID group. Quantitative analysis confirmed that the three systems could accurately distinguish between the two secretory pathways without cross-interference, providing a specificity basis for subsequent in vivo experiments.
Figure 5 | Biotinylation of Secreted Peptides in Cell Culture
✦ (2) Dynamic Remodeling of the Hepatocyte Secretome
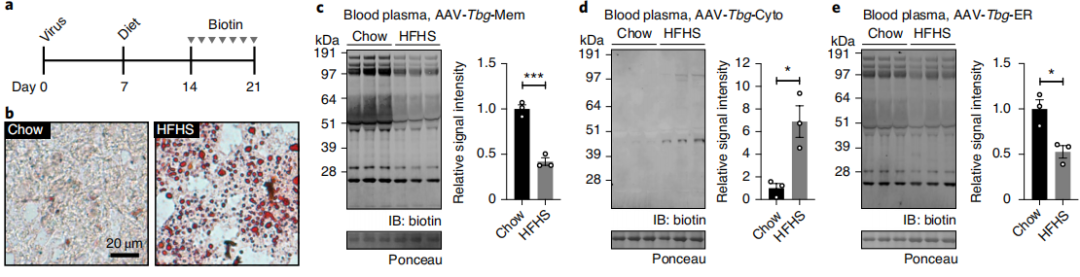
Cellular secretory function undergoes dynamic changes in response to environmental stress and physiological perturbations. A high-fructose high-sucrose (HFHS) diet was used to establish a model of hepatic lipid accumulation in order to investigate the regulation of the hepatocyte secretome under metabolic stress. Mice received tail vein injection of hepatocyte-specific AAV-TBG vectors (AAV8-TBG-ER-TurboID, AAV8-TBG-Cyto-TurboID, or AAV8-TBG-Mem-TurboID). Seven days after transduction, the mice were divided into a normal diet group and an HFHS diet group and maintained under the respective dietary conditions for 2 weeks. From Day 14 to Day 21, biotin was administered by intraperitoneal injection to complete in vivo labeling. Samples were collected 24 hours after the final administration. Oil Red O staining confirmed that the HFHS diet markedly induced hepatic lipid accumulation. Immunoblot analysis of plasma proteins showed that, under HFHS-induced metabolic stress, the abundance of classical secretory proteins labeled by ER-TurboID and Mem-TurboID decreased by 50%–60%, whereas unconventional secretion detected by Cyto-TurboID increased approximately 7-fold, with the signal predominantly concentrated in a 45-kDa protein band. Intracellular control experiments demonstrated no difference in protein biotinylation levels within hepatocytes between the two groups, indicating that the changes in plasma signals resulted from remodeling of secretory pathways rather than alterations in labeling efficiency. This system enables sensitive detection of dynamic changes in the in vivo secretome under physiological and pathological perturbations.
Figure 6 | Dynamic and Nutrient-Dependent Remodeling of the Hepatocyte Secretome
IV. Quick Selection Guide
✅ Neuron–astrocyte interactions and proteins at cell–cell contact sites → Split-TurboID
✅ Whole-brain astrocyte networks and gap junction function → Cx43-TurboID
✅ Secretome profiling, plasma protein source tracing, and metabolic perturbation studies → Cyto-TurboID / Mem-TurboID / ER-TurboID
V. Shared Technical Advantages
💡1. Low toxicity and compatible with in vivo applications. Only exogenous biotin is required, ensuring safety for long-term animal studies.
💡2. Covalent labeling. Biotin labeling remains stable throughout fixation, tissue clearing, and cell lysis, preventing signal loss during sample processing.
💡3. Compatible with both mass spectrometry-based proteomics and fluorescence imaging. The labeled samples can be analyzed by either downstream approach, making the system adaptable to multidisciplinary applications.
💡4. Cell type-specific targeting enabled by AAV. AAV-mediated delivery enables cell type-specific expression, minimizing background signals from non-target cell populations.
References
Branon TC, Bosch JA, Sanchez AD, et al. Efficient proximity labeling in living cells and organisms with TurboID. Nat Biotechnol. 2018;36(9):880–887.
Takano T, Wallace JT, Baldwin KT, et al. Chemico-genetic discovery of astrocytic control of inhibition in vivo. Nature. 2020;588(7837):296–302.
Cooper ML, Selles MC, Cammer M, et al. Astrocytes connect specific brain regions through plastic networks. Nature. Published online April 22, 2026.
Wei W, Riley NM, Yang AC, et al. Cell type-selective secretome profiling in vivo. Nat Chem Biol. 2021;17(3):326–334.
Service Type :
Select the service you'd like to purchase.
Order Information(Premade-AAVs)
Please provide us some information about the service you'd like to order.
Order Information(Custom AAV/Lentivirus)
Please provide us some information about the service you'd like to order.
Order Information(Others)
Please provide us some information about the service you'd like to order.