The world’s first red fluorescent potassium sensors,RGEPO1/2,are here! |A new tool for deciphering neurological diseases—with expert commentary from Prof. Yulong Li’s team at Peking University,revealing the technical limitations and future direction
Time:2025-11-06 09:14:47
The homeostatic balance and dynamic fluctuations of ions are crucial for a wide range of biological processes, including neuronal signal transmission and cellular homeostasis. Detecting these ion changes under physiological conditions is essential for elucidating their functional roles in health and disease. Traditional detection methods, however, are limited by invasiveness and low resolution, making them insufficient for advanced research needs. Genetically encoded potassium indicators (GEPOs) offer a promising alternative, enabling real-time monitoring of ion concentration changes with minimal invasiveness and high spatial and temporal resolution.
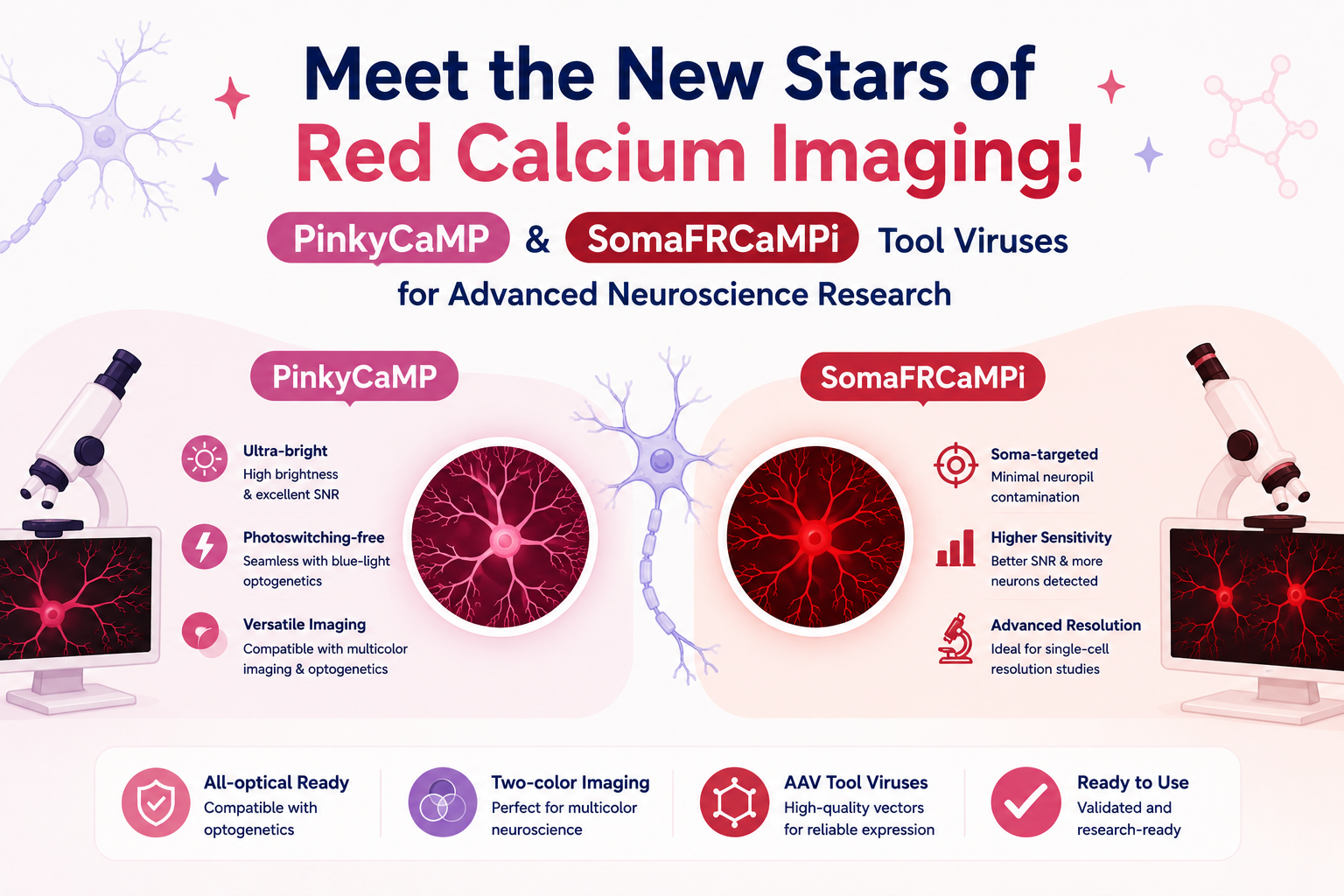
Most existing calcium and potassium indicators emit green fluorescence, but red indicators offer key advantages — lower phototoxicity, deeper tissue penetration, and compatibility for multi-color imaging with other indicators. However, reported red fluorescent calcium indicators have shown suboptimal sensitivity and performance. On February 2, 2025, Kiryl D. Piatkevich’s team at Westlake University published a paper in PLOS Biology titled “A Sensitive Soma-localized Red Fluorescent Calcium Indicator for Multi-Modality Imaging of Neuronal Populations In Vivo.” The team developed two improved calcium indicators,FRCaMPi and its soma-localized version SomaFRCaMPi, with the latter performing comparably to the best green soma-localized GECIs.
Compared with red calcium indicators, the development of red potassium indicators had long remained a challenge. On September 17, 2025, Kiryl D. Piatkevich’s team at Westlake Laboratory, in collaboration with Alice R. Walker’s group at Wayne State University, published an article in PLOS Biology titled “Sensitive red fluorescent indicators for real-time visualization of potassium ion dynamics in vivo.” Through directed evolution in E. coli and further optimization in mammalian cells, the researchers developed two red genetically encoded potassium indicators, RGEPO1 and RGEPO2. These sensors enabled, for the first time, the visualization of intracellular and extracellular potassium transients during epileptic seizures in mice, providing a powerful new tool for understanding the role of potassium ions in neuronal activity and disease.
Development and Validation of Red Fluorescent Potassium Indicators
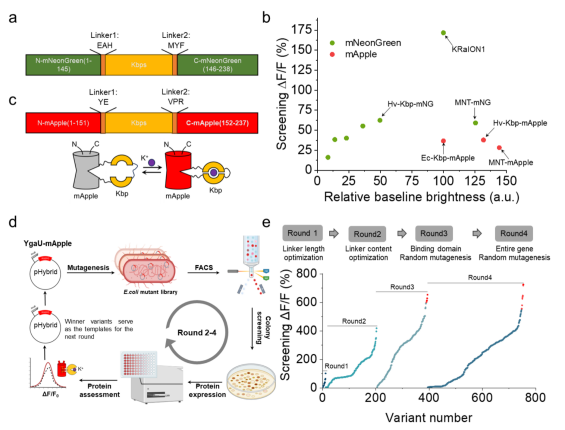
To develop a single red fluorescent protein-based potassium ion indicator, the researchers used the known K⁺-binding protein Hv-Kbp as a template. Through genome mining and NCBI BLAST searches, they identified five homologous proteins containing conserved potassium-binding sites. Each homolog was substituted for Ec-Kbp in the KRaION1 indicator and expressed in E. coli for validation (Fig. 1a). Based on combined brightness and response amplitude, Hv-Kbp was ultimately selected and inserted into the red fluorescent protein mApple, forming the initial sensor prototype (Fig. 1b–d).
Using high-throughput screening in E. coli, the team applied random mutagenesis and optimized linker length and sequence, obtaining the best-performing variant with a maximum ΔF/F exceeding 600% (Fig. 1e). However, the sensor’s in vitro performance did not directly translate to mammalian cells — the optimal variant showed only 1/13 of the fluorescence response observed in solution when expressed in HEK293FT cells, suggesting that intracellular environmental factors may alter protein conformation and potassium responsiveness (Fig. 2a–b).
Figure 1. Optimization of red potassium indicators in E. coli
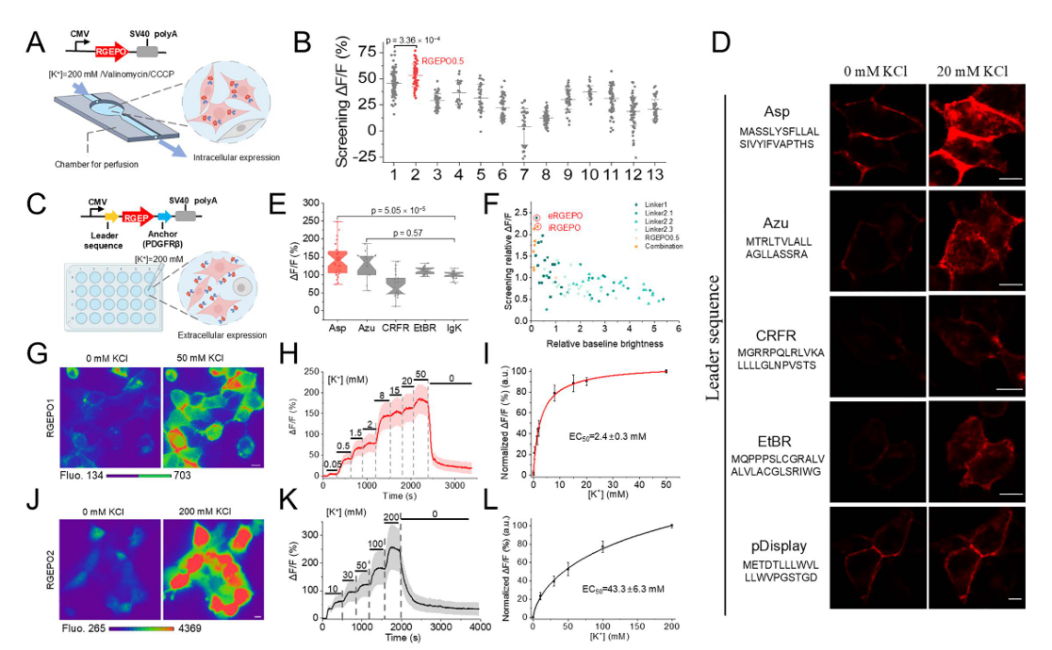
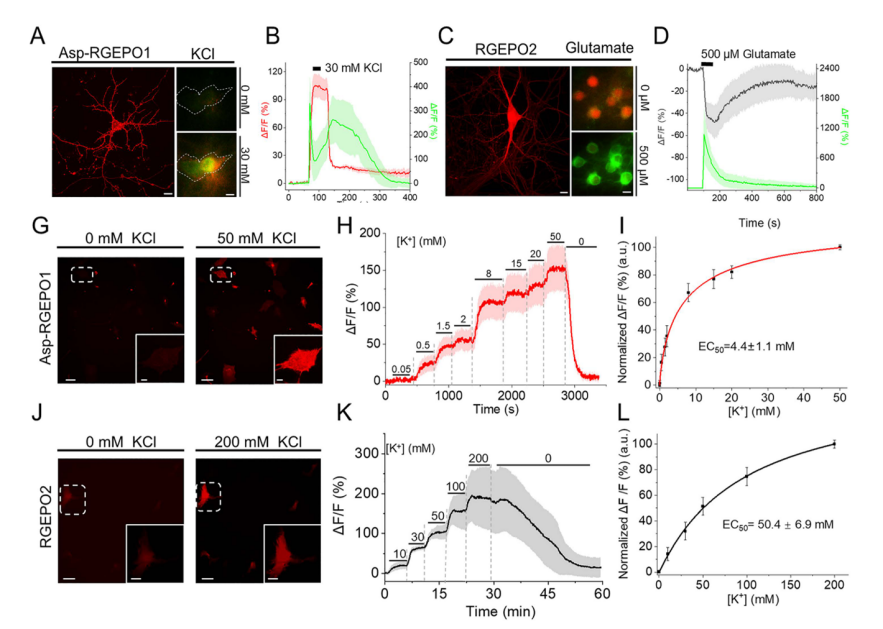
To address the performance drop caused by the cellular environment, the team fused RGEPO0.5 with various N-terminal signal peptides and a PDGFRβ transmembrane domain to target the probe to the cell surface, facilitating rapid buffer exchange and enhancing screening efficiency. Among these, the aspartate signal peptide variant exhibited the best performance (Fig. 2c–e). Using this Asp–PDGFRβ surface-display system, they conducted saturation mutagenesis of the linker region, identifying two top-performing variants: RGEPO1, which showed the strongest extracellular K⁺ response, andRGEPO2, which showed the strongest cytosolic K⁺ response,with their primary difference lying in the linker region sequences (Fig. 2f).
Titration assays in HEK293FT cells revealed distinct localization and sensitivity profiles: RGEPO1 was membrane-localized, responsive to extracellular K⁺, with an EC₅₀ of 2.4 ± 0.3 mM and a dynamic range of 0.5–20 mM (Fig. 2g–i). RGEPO2 was cytosolic and nuclear, responsive to intracellular K⁺, with an EC₅₀ of 43.3 ± 6.3 mM and a detection range of 4–200 mM. Both sensors exhibited reversible fluorescence changes upon K⁺ fluctuation (Fig. 2j–l). Together, RGEPO1 and RGEPO2 represent two optimized red fluorescent potassium indicators derived from the novel Hv-Kbp domain, capable of monitoring intracellular and extracellular K⁺ dynamics across a broad range of 0.5–200 mM.
Figure 2. Optimization process of red potassium indicators in HEK cells
Characterization of RGEPOs in Solution
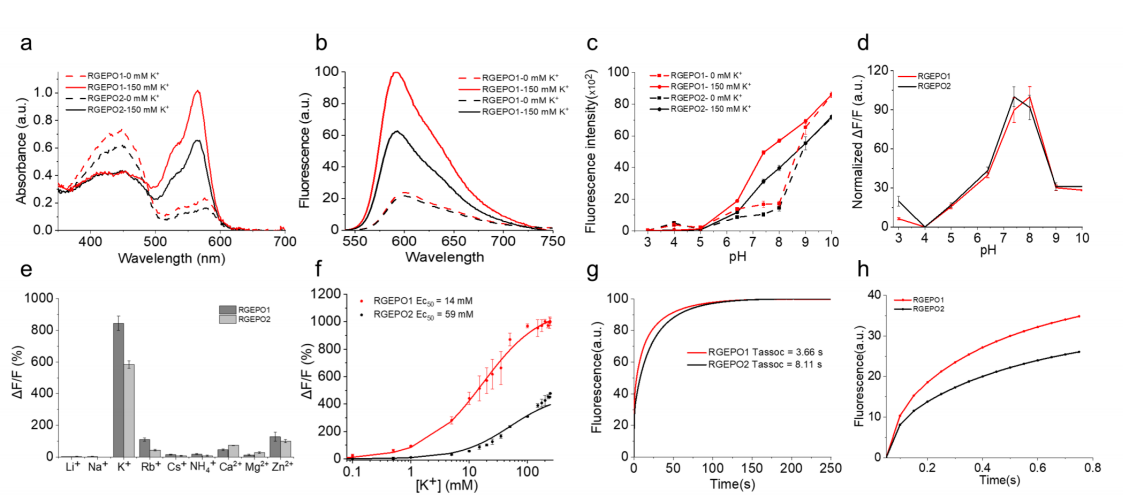
To evaluate the suitability of RGEPOs for use in mammalian systems, the researchers characterized RGEPO1 and RGEPO2 spectrally across the physiological potassium concentration range (0–150 mM, pH 7.4). Both sensors exhibited dual absorbance peaks in the absence of K⁺ — at 448/446 nm (protonated) and 576/578 nm (deprotonated). Upon K⁺ addition, the deprotonated peak increased while the protonated peak decreased. The excitation and emission maxima were 575/574 nm and 591/593 nm, respectively, with fluorescence intensity increasing 4.7-fold for RGEPO1 and 3.1-fold for RGEPO2 (Fig. 3a–b). These changes resulted from enhanced extinction coefficients and quantum yields. Both sensors also exhibited potassium-dependent fluorescence under two-photon excitation, indicating their suitability for in vivo imaging applications.
To further assess pH sensitivity and ion selectivity, both indicators demonstrated marked fluorescence responses to K⁺ within the pH 6.5–9.0 range, with maximal dynamic ranges observed at pH 8.0 (RGEPO1) and pH 7.4 (RGEPO2) (Fig. 3c–d). Testing against various metal ions revealed that only K⁺ induced the highest ΔF/F changes — 844% for RGEPO1 and 584% for RGEPO2 — and neither responded to Ca²⁺, confirming their high selectivity for potassium (Fig. 3e).
Kinetic analysis showed that the half-time to reach plateau fluorescence upon K⁺ binding was approximately 3.66 s for RGEPO1 and 8.11 s for RGEPO2 (Fig. 3g–h). Under isotonic conditions, the EC₅₀ values were 14 mM (RGEPO1) and 59 mM (RGEPO2), with an overall dynamic range of 0.1–225 mM (Fig. 3f).
Collectively, the fluorescence spectral characterization, brightness, and kinetic properties demonstrate that RGEPO1 and RGEPO2 exhibit high selectivity and sensitive responses to potassium ions across different physiological environments, making them ideal indicators for monitoring potassium ion dynamics.
Figure 3. In vitro characterization of RGEPOs
Molecular Mechanism Underlying the Potassium Sensitivity of RGEPOs
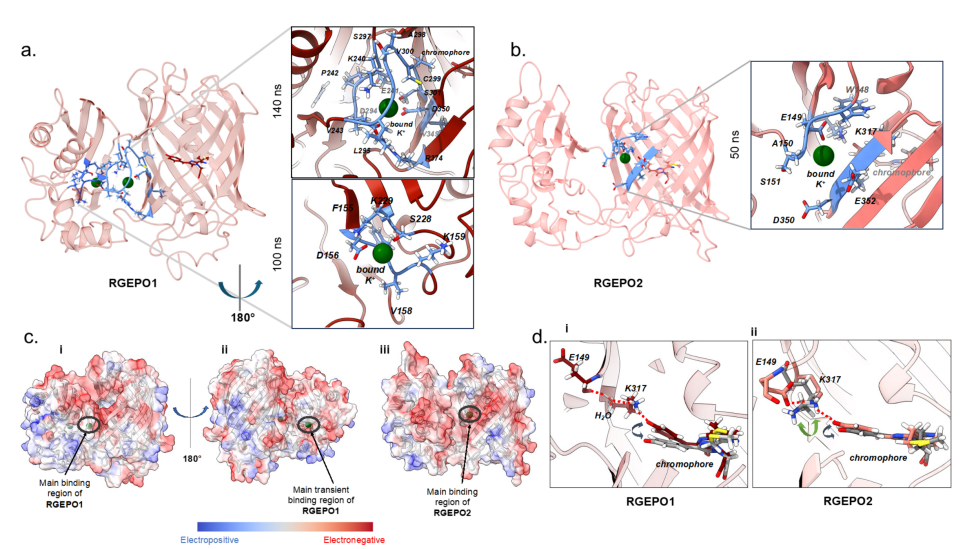
Molecular dynamics simulations of RGEPO1 revealed the presence of two potassium-binding pockets — a primary binding site and a transient binding site. The primary pocket comprises 13 amino acid residues, including D294 and D350, which directly coordinate with K⁺, and R374, which stabilizes the ion through a hydrogen bond with D294 (Fig. 4a). A continuous electronegative surface extends from the primary to the transient binding site, favoring the attraction and prolonged retention of positively charged ions (Fig. 4c).
In contrast, RGEPO2 contains only one binding pocket composed of seven amino acids, located near the fluorescent protein domain. Upon K⁺ binding, a series of hydrogen-bond interactions form between the chromophore phenolate group and K317, triggering changes in the fluorescence signal (Fig. 4b). The pocket is also surrounded by negatively charged residues, which facilitate stable cation binding (Fig. 4c).
Molecular dynamics analyses confirmed that both sensors share the same fluorescence-sensing mechanism: potassium binding to the primary binding pocket induces hydrogen bond interactions with the O-position of the chromophore phenolate ring. The key difference lies in the mediation of this interaction — RGEPO1 requires solvent mediation, whereas RGEPO2 does not (Fig. 4d). Taken together, these results indicate that the primary potassium-binding pocket in both sensors is closely associated with their fluorescence response mechanism.
Figure 4. Molecular mechanism underlying the potassium sensitivity of RGEPOs
RGEPOs Record Dynamic Potassium Ion Changes in Primary Neurons and Astrocytes
When RGEPO1 was expressed on the extracellular side of the plasma membrane and GCaMP6f in the cytoplasm of primary hippocampal neurons (Fig. 5a), stimulation with 30 mM K⁺ led to fluorescence increases of approximately 71% (RGEPO1) and 350% (GCaMP6f), with peak times of 27 s and 3s, respectively, both returning to baseline after washout (Fig. 5b). Since excessive glutamate stimulation of glutamate receptors is known to cause massive K⁺ efflux from neurons, co-expression of RGEPO2 (evenly distributed in neuronal soma and dendrites) and GCaMP6f in the same hippocampal neurons showed that after 500 µM glutamate stimulation, GCaMP6f fluorescence increased by ~1076% with a peak at 9 s, while RGEPO2 fluorescence decreased by ~48% with a peak at 72 s (Fig. 5d). These results demonstrate that RGEPOs can effectively monitor transient intra- and extracellular K⁺ dynamics in neurons.
Compared with neurons, astrocytes play a crucial role in regulating and maintaining stable extracellular potassium levels, which are essential for central nervous system homeostasis. In primary mouse astrocytes, RGEPO1 fluorescence increased with rising extracellular K⁺ concentrations, reaching a ΔF/F of 206% at 50 mM and exhibiting a surface dissociation constant (Kd) of 4.4 ± 1.1 mM. Meanwhile, cytosolic RGEPO2 fluorescence also depended on K⁺ concentration, showing a ΔF/F of 328% at 200 mM with an EC₅₀ of 50 ± 6.9 mM (Fig. 5g–l). These findings indicate that elevated extracellular K⁺ triggers potassium uptake by astrocytes, supporting their role in K⁺ spatial buffering. Under similar depolarizing conditions, neurons exhibited distinct potassium responses, highlighting cell-type-specific K⁺ dynamics.
Figure 5. RGEPOs record dynamic potassium ion changes in primary neurons and astrocytes
RGEPOs Record Transient Potassium Ion Changes in Neurons of Acute Brain Slices
Using recombinant adeno-associated virus (rAAV), the researchers co-expressed RGEPO1 or RGEPO2 together with GCaMP6f in cortical neurons of neonatal mice. At 3–4 weeks post-infection, immunohistochemistry confirmed the expression of RGEPOs in cortical regions (Fig. 6a–c). After 6 weeks of expression, acute brain slices were prepared for functional imaging.
Upon stimulation with 20 or 30 mM KCl, RGEPO1 fluorescence increased by 6.5%, while RGEPO2 fluorescence decreased by 7.0%; simultaneously, GCaMP6f fluorescence increased by 104% and 128%, respectively. In all cases, fluorescence signals returned to baseline after washout (Fig. 6d–i).
Although the ΔF/F values for potassium detection in acute brain slices were smaller than those in cultured neurons — likely due to differences in protein expression levels and tissue regulatory networks — the overall response trends were consistent. These findings demonstrate that RGEPOs can effectively monitor intra- and extracellular potassium dynamics in intact brain tissue.
Figure 6. RGEPOs record transient potassium ion changes in neurons of acute brain slices
RGEPOs record dynamic potassium changes during epileptic seizures in mice
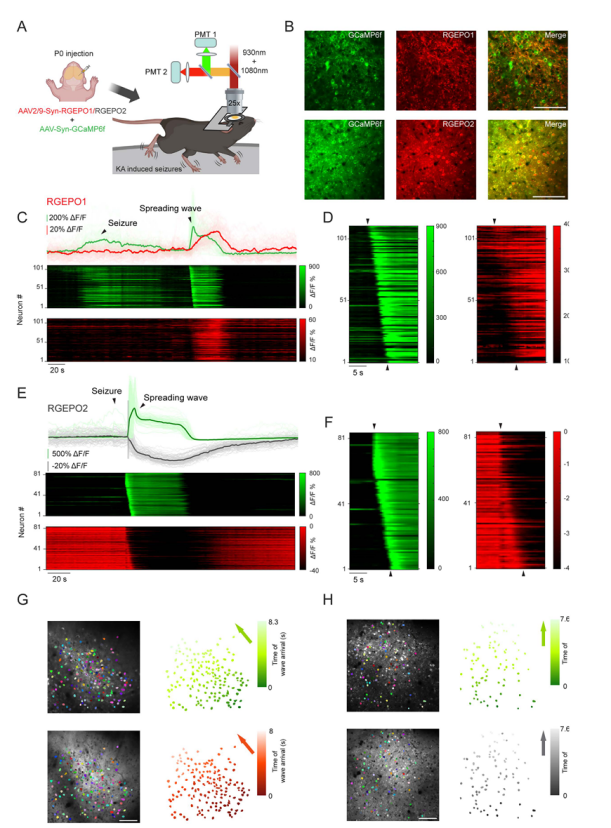
Epilepsy is a severe neurological disorder characterized by defects in potassium channels and disrupted potassium homeostasis, leading to brain hyperexcitability and seizure activity. In this study, the research team co-expressed GCaMP6f and RGEPOs in neurons of the primary somatosensory cortex, and used two-photon microscopy to monitor L2/3 neuronal activity during seizures induced by kainic acid (KA) (Fig. 7a–b).
Consistent with previous findings, calcium imaging revealed synchronous fluorescent waves followed by global propagating waves, corresponding to the ictal (seizure) phase and the spreading depolarization phase, respectively (Fig. 7c–e).
After seizure termination, RGEPO1 showed a fluorescence increase with a median peak ΔF/F of 89%, forming a propagating wave spatially matched with GCaMP6f signals but with a broader front (Fig. 7c,d,g). This may result from RGEPO1’s slower kinetics, producing a smoother response.
In contrast, RGEPO2 exhibited a rapid fluorescence decrease (median peak ΔF/F = –52%) synchronized with GCaMP6f and followed by a propagating wave, indicating a drop in intracellular potassium levels (Fig. 7e,f,h).
These findings demonstrate that despite slower kinetics, RGEPO1 and RGEPO2 reliably report potassium concentration dynamics in vivo during epileptic seizures in mice, achieving the first single-cell–resolution observation of potassium kinetics at the population level in a drug-induced epilepsy model, while remaining compatible with the calcium indicator GCaMP6f.
Figure 7. Two-photon imaging of GCaMP6f and RGEPOs in the kainic acid–induced mouse epilepsy model.
Summary
RGEPOs are the first red genetically encoded potassium indicators, enabling real-time visualization of intracellular and extracellular potassium dynamics across various biological systems, including cell cultures, brain slices, and live animals. Although RGEPOs exhibit lower temporal and signal amplitude resolution compared with calcium indicators such as GCaMP6f, this study provides a novel tool for investigating the roles of potassium in physiological and pathological processes and paves the way for further development of red fluorescent protein-based biosensors.
Expert Commentary
👨🏫Yulong Li, School of Life Sciences, Peking University
The two studies by Kiryl D. Piatkevich’s team at Westlake University represent significant breakthroughs in the field of red-shifted ion imaging. They not only developed optimized red calcium indicators (FRCaMPi and SomaFRCaMPi) but also, for the first time, successfully created red potassium indicators (RGEPO1 and RGEPO2), greatly expanding the toolkit available for imaging neuronal and other tissue cellular activities.
These red indicators offer advantages such as deep tissue penetration, multi-channel monitoring, and low invasiveness, providing a novel technical approach for dissecting neural dynamics and ion signaling across different biological systems. They can be efficiently combined with green fluorescent indicators to achieve simultaneous imaging of calcium/potassium dynamics and neurotransmitter or neuromodulator signals. This combination allows researchers to track neuronal activity alongside neurochemical events — such as the release of small-molecule neurotransmitters or neuropeptides — in live animals under physiological or pathological conditions, providing critical data for understanding neural regulation across brain regions.
A current limitation of this technology is that indicators based on mApple (e.g., jRGECO1a) are susceptible to blue-light–induced photoactivation, which restricts their compatibility with blue-light-driven optogenetic tools. Future research should focus on reducing photoactivation sensitivity while improving photostability, ion selectivity, and dynamic range to fully realize the potential of these indicators in complex experimental settings.