Client Publication | Neuron | Liping Wang and Xuemei Liu’s Team at SIAT Uncovers Neural Mechanisms Behind Individual Differences in Innate Fear Responses
Release time:2025-09-24 11:59:41
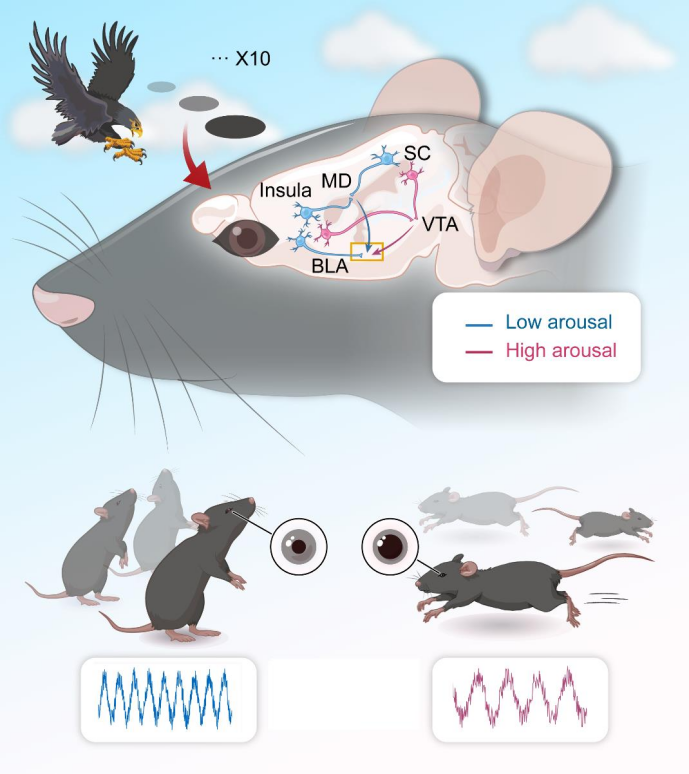
Emotional responses, particularly fear-related behaviors, are critical for survival when facing threats. When exposed repeatedly to predator threats, animals exhibit two distinct adaptive strategies—habituation and sensitization—but the underlying neural circuit mechanisms remain unclear. Looming visual stimuli (LS) can trigger innate defensive responses (T1) and habituated behaviors (T2), yet the circuits regulating arousal levels and habituation are not fully understood. On May 9, 2025, a research team led by Prof. Liping Wang and Associate Prof. Xuemei Liu at the Institute of Brain Cognition and Brain Disease, Shenzhen Institute of Advanced Technology (SIAT), Chinese Academy of Sciences / Shenzhen-Hong Kong Institute of Brain Science, published a paper in Neuron titled “Neural Circuit Underlying Individual Differences in Visual Escape Habituation.” The study identified two behavioral types in response to repeated visual threats and mapped their corresponding neural circuits: The SC-VTA-BLA pathway (Superior Colliculus → Ventral Tegmental Area → Basolateral Amygdala) mediates T1 responses. The SC-MD-BLA and Insula-MD-BLA pathways (SC/Insula → Mediodorsal Thalamus → BLA) mediate T2 responses. The MD functions as a central hub that integrates input from both the SC and the insular cortex, modulating arousal and defensive behaviors.
Prof. Liping Wang is the paper’s senior corresponding author, and Associate Prof. Xuemei Liu is both first author and co-corresponding author. Dr. Juan Lai (Assistant Researcher at SIAT) and Dr. Chuanliang Han (now a Postdoctoral Fellow at The Chinese University of Hong Kong) are co-first authors. The research received significant support from SIAT Brain Institute researchers Prof. Fuqiang Xu, Prof. Liming Tan, and Prof. Pengfei Wei.
Individual Differences in Escape Habituation to Repeated Visual Threat Stimuli
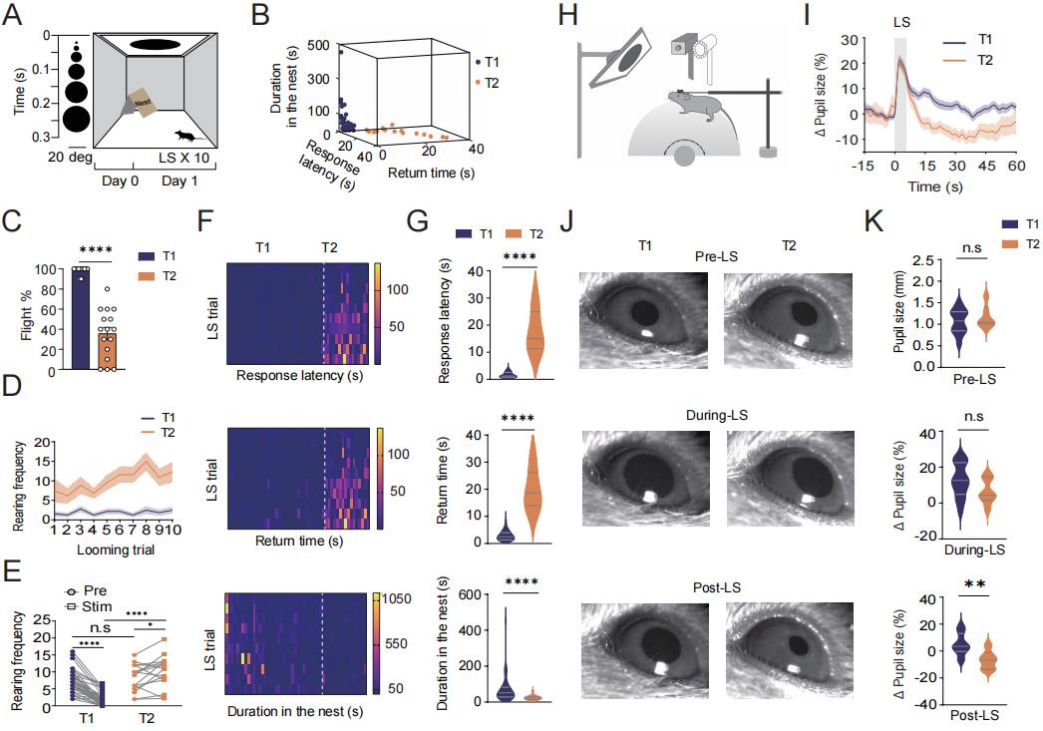
Previous studies have confirmed that LS reliably trigger innate escape behaviors. To investigate the phenomenon of escape habituation—defined as the gradual reduction of escape responses to repeated threat stimuli—52 wild-type adult male mice were subjected to 10 LS trials. Each trial lasted 5.5 seconds, with inter-trial intervals of at least 2 minutes (Figure 1A). Based on average response latency, the mice were categorized into two groups: "persistent escapers" (T1) and "rapid habituators" (T2). The T1 group had average response latencies shorter than 5.5 seconds, while those in the T2 group exceeded this threshold (Figure 1B).
Compared to T1 mice, T2 mice exhibited a significantly lower average escape ratio (Figure 1C), longer response latency and return-to-nest time, and shorter nest stay duration (Figures 1F–G). During the trials, T2 mice also showed a stage-dependent increase in both latency and return time. In terms of non-selective attention—measured via rearing frequency—T2 mice displayed a higher baseline level (Figure 1D) and a significant post-stimulus increase, whereas the T1 group showed a decrease (Figure 1E).
Regarding arousal state, 10 seconds after LS exposure, the pupil diameter in the T1 group was significantly larger than in the T2 group, while there was no notable difference between the two groups before or during the stimulus (Figures 1I–K). These findings suggest that T2 mice demonstrate a higher level of habituation to repeated LS exposure, characterized by increased stimulus-evoked non-selective attention and decreased arousal.
Figure 1. Individual Differences in Escape Habituation to Repeated Looming Stimuli
Distinct Superior Colliculus (SC) Pathways Regulate Arousal and Escape Behaviors
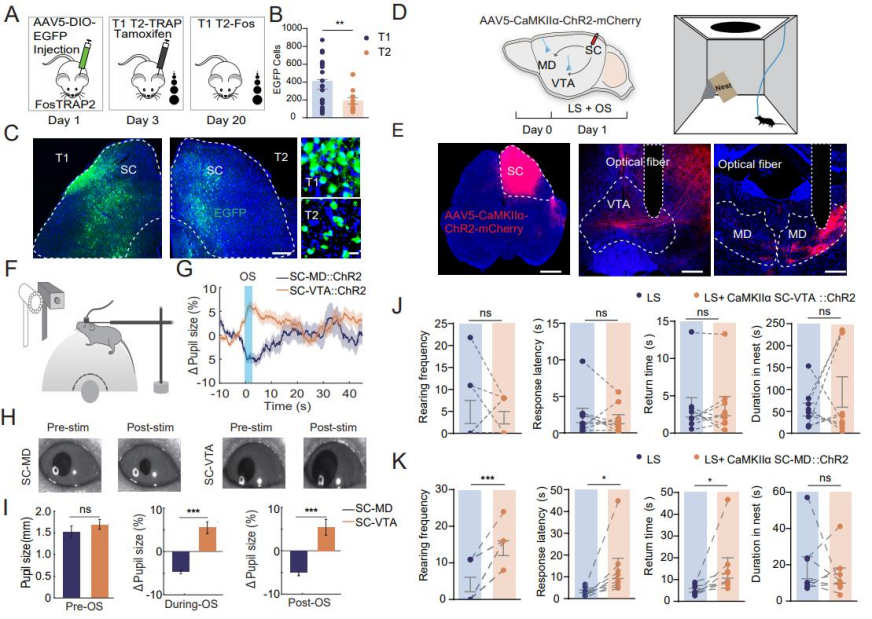
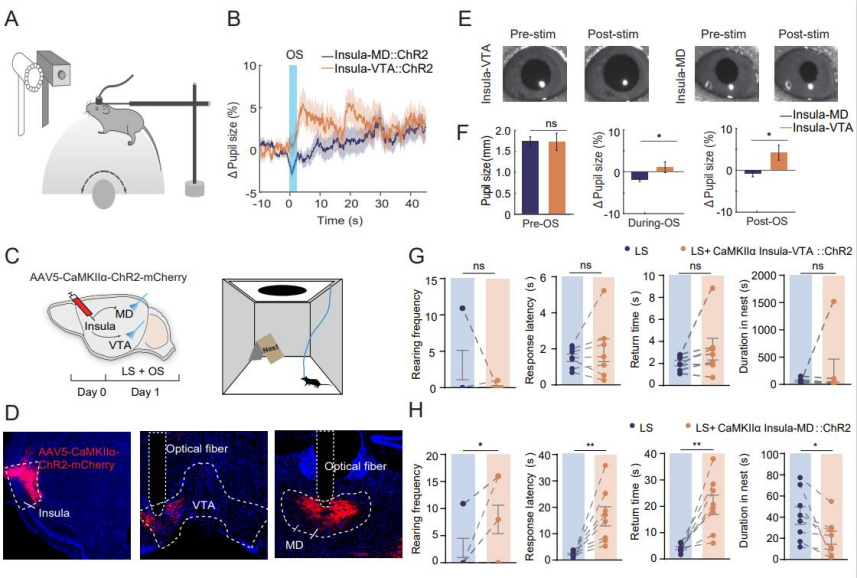
To clarify the role of SC neurons activated during LS habituation, researchers employed the FosTRAP2 technique. AAV5-DIO-EGFP was unilaterally injected into the SC of FosTRAP2 mice. On day 3, tamoxifen was administered, and the mice were exposed to repeated LS stimulation to capture activated cells. On day 24, mice were re-exposed to LS prior to perfusion (Figure 2A). Results showed a higher density of EGFP⁺ neurons in the SC of T1 mice compared to T2 mice, indicating that SC activity is involved in the LS habituation process and contributes to the attenuation of innate escape responses (Figures 2B–C). To investigate whether the SC–VTA and SC–MD pathways differentially influence arousal during LS habituation, AAV5-CaMKIIα-ChR2-mCherry was injected into the SC, and optical fibers were implanted in the MD and VTA (Figures 2D–E). Upon LS exposure: Activation of the SC–MD pathway led to pupil constriction. Activation of the SC–VTA pathway led to pupil dilation (Figures 2F–G). During and after light stimulation, the average pupil diameter in the SC–VTA group was significantly larger than in the SC–MD group (Figures 2H–I), demonstrating distinct effects of the two pathways on arousal levels.
Repeated SC–VTA stimulation in an open field with cover induced stable escape behaviors similar to those seen in the T1 group. In contrast, SC–MD stimulation increased rearing frequency, response latency, and return time—disrupting escape behavior. SC–VTA stimulation had no such disruptive effects. These findings suggest: SC–MD activation lowers arousal, increases non-selective attention, and impairs innate escape behavior. SC–VTA activation elevates arousal without affecting non-selective attention or escape behavior.
Figure 2. Distinct SC Pathways Regulate Arousal and Escape Behavior
SC Neurons Projecting to VTA and MD Functionally Converge on the BLA
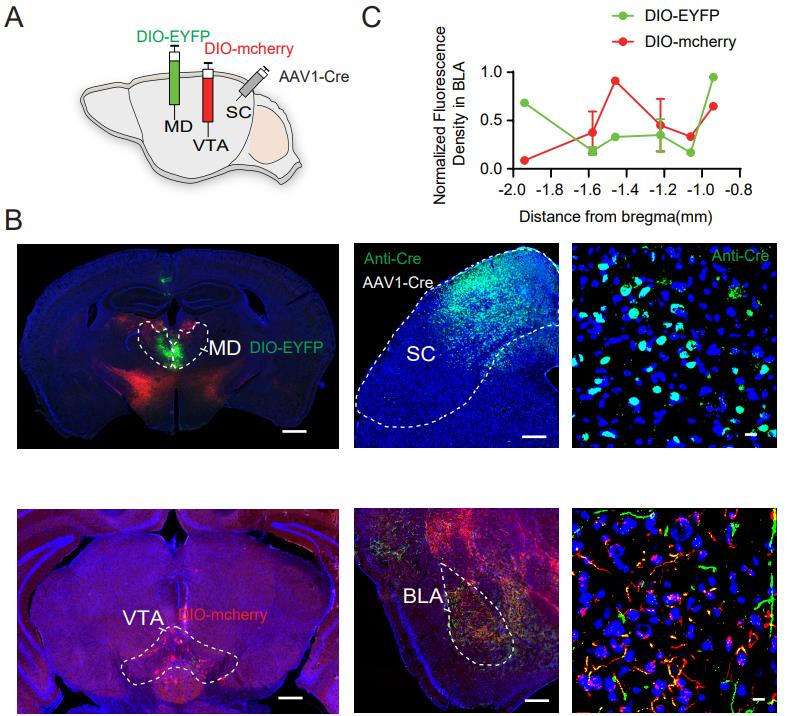
The amygdala plays a critical role in both conditioned and innate fear responses, with the BLA serving as a central hub for encoding threat-related information. The authors investigated whether SC neurons projecting to the VTA and the MDalso converge onto the BLA. To test this, AAV1-Cre was injected into the SC, AAV5-DIO-EYFP into the MD, and AAV5-DIO-mCherry into the VTA (Figure 3A). The results revealed overlapping axonal projections within the BLA, indicating that SC neurons projecting to both the VTA and MD target a shared region within the BLA (Figure 3B). Fluorescence density analysis further showed that: SC neurons projecting to the VTA primarily innervate the medial part of the BLA. SC neurons projecting to the MD mainly target the anterior and posterior regions of the BLA (Figure 3C).
Figure 3. SC Neurons Projecting to VTA and MD Converge on the BLA
SC Pathways Regulate Connectivity and Oscillatory Activity in the BLA
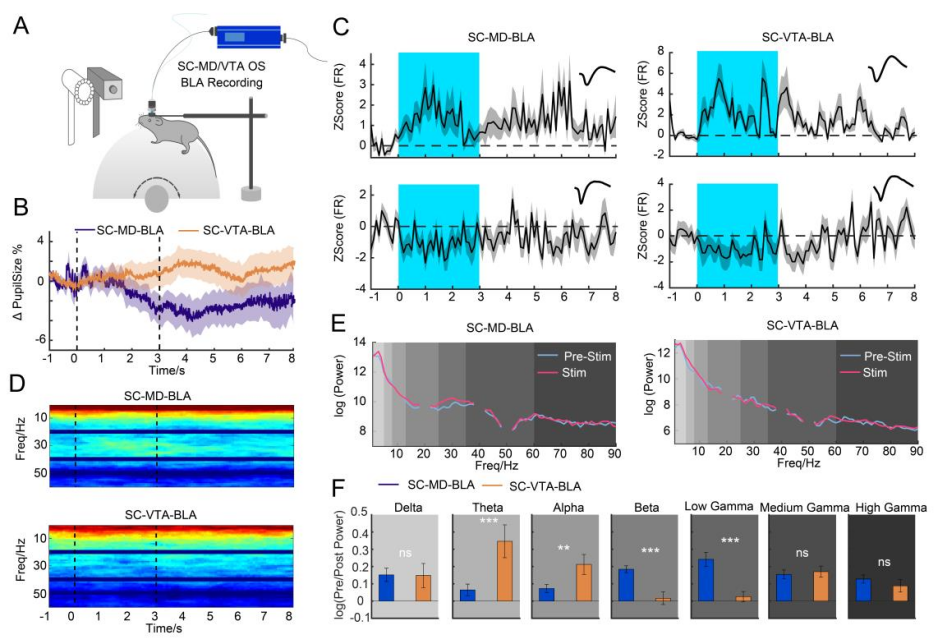
To further investigate the functional connectivity of SC pathways, researchers selectively activated the SC–VTA and SC–MD circuits using optogenetics while simultaneously recording in vivo multi-channel activity and pupil dynamics in the BLA (Figure 4A). Activation of the SC–VTA pathway resulted in pupil dilation, while SC–MD activation led to pupil constriction (Figure 4B). Following pathway activation, significant changes were observed in the firing rates of BLA neurons. Power spectral analysis of local field potentials (LFPs) in the BLA revealed distinct oscillatory patterns associated with each pathway. (Oscillations refer to rhythmic fluctuations in neuronal activity over time, measurable through LFPs. They reflect coordinated activity across neural populations, with specific frequency bands linked to distinct neural functions and behaviors.) SC–VTA activation increased power in low-frequency bands (theta and alpha). SC–MD activation enhanced power in higher-frequency bands (beta and low gamma) (Figures 4D–F). These findings demonstrate that the SC–MD and SC–VTA pathways regulate the BLA through distinct oscillatory dynamics, underscoring their unique roles in amygdala circuitry and fear-related behavior.
Figure 4. SC Pathways Regulate BLA Connectivity and Oscillatory Patterns
Distinct Modulatory Roles of Insular Cortex Pathways in Arousal and Defensive Behaviors
The insular cortex is a key integrative hub for sensory feedback and autonomic arousal, known to project to both the MD and the VTA. To examine the functional roles of these projections, AAV5-CaMKIIα-ChR2-mCherry was injected into the insular cortex, and optical fibers were implanted in the MD and VTA (Figures 5A, 5D). Optogenetic stimulation combined with pupilometry revealed: Stimulation of insula–MD terminals caused pupil constriction. Stimulation of insula–VTA terminals led to pupil dilation (Figures 5E–F). These results indicate that the insula–MD and insula–VTA pathways modulate arousal in distinct ways. In mice expressing ChR2-mCherry in insular neurons, insula–MD activation increased rearing frequency, response latency, and return time, while decreasing time spent in the nest—indicative of reduced arousal, enhanced non-selective attention, and weakened LS-induced escape behavior. In contrast, insula–VTA activation elevated arousal but did not affect LS-evoked behaviors (Figures 5C–D, 5G–H).
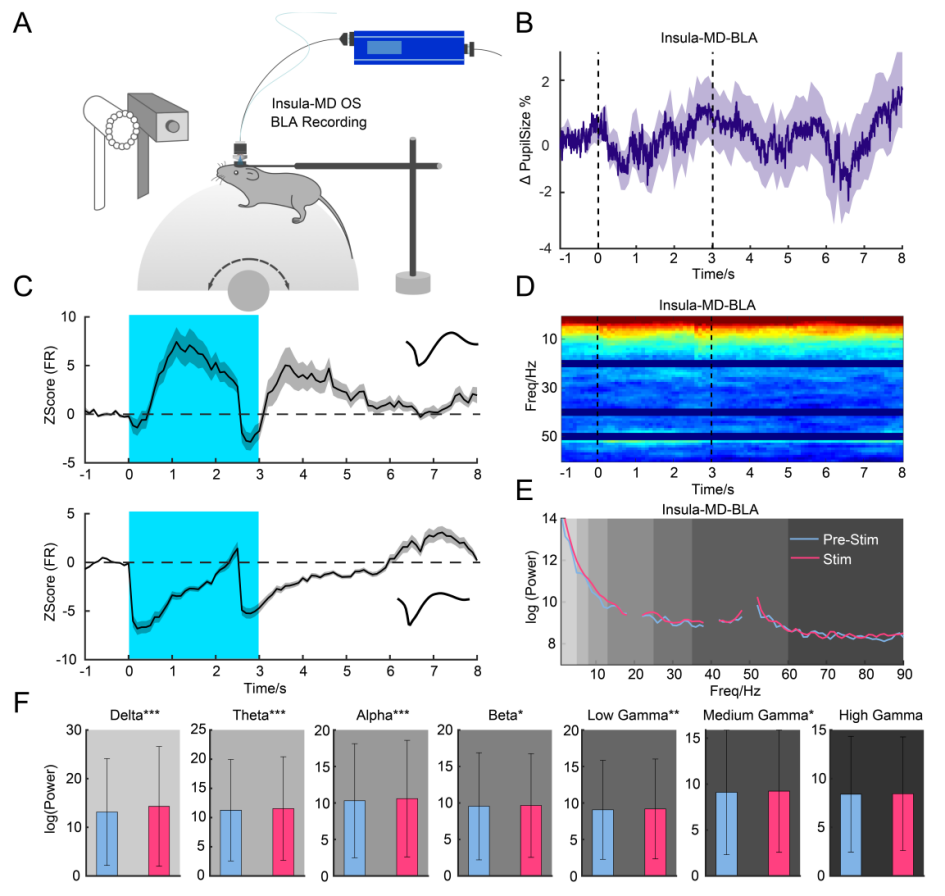
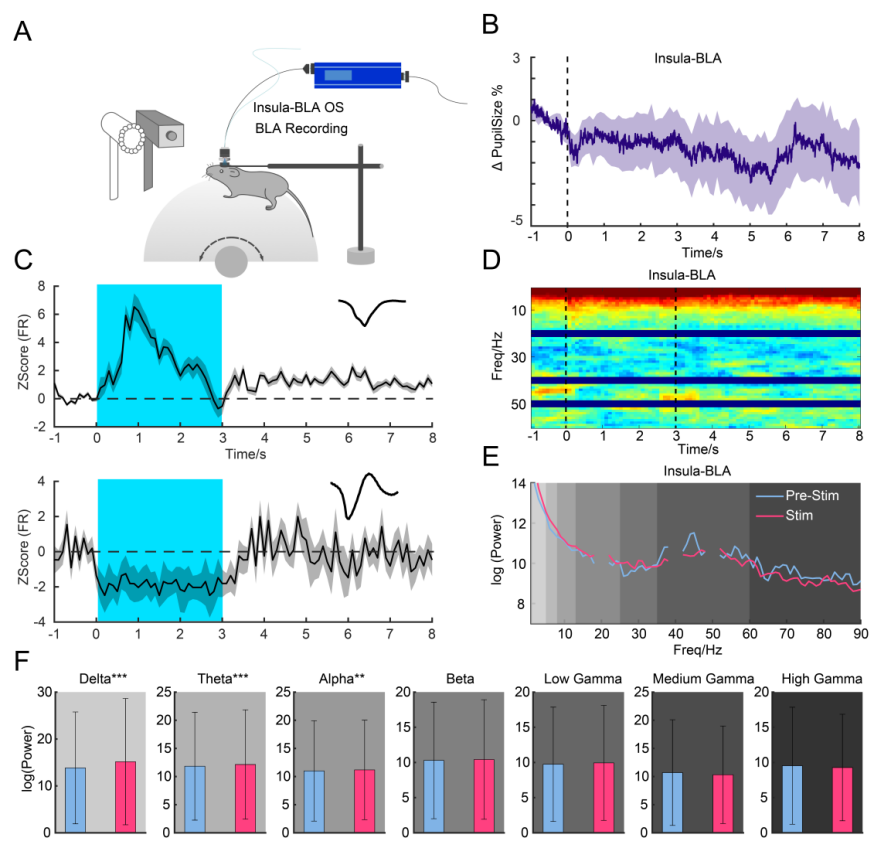
To further assess functional connectivity, multi-channel in vivo recordings and pupilometry were conducted during insula–MD pathway activation (Figure 6A). Stimulation of this pathway led to pupil constriction and altered neuronal firing in the BLA, with 30% of neurons responding (Figure 6B). However, no significant changes were observed in LFP power across frequency bands (Figures 6D–F).
Figure 6. Insula–MD Pathway Modulates BLA Activity
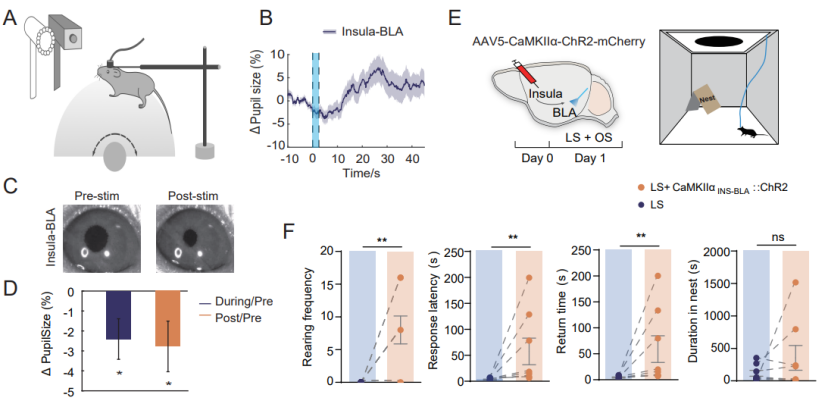
Using optogenetic manipulation and pupil measurement to investigate whether the insula–BLA pathway affects arousal (Figure 7). After injecting ChR2-mCherry into the insular cortex and implanting optical fibers in the BLA, optogenetic stimulation reduced pupil size (Figures 7B–D). Activation of the insula–BLA pathway increased rearing frequency, response latency, and return time, indicating a reduction in LS-induced escape behavior.
Figure 7. Effects of the insula–BLA pathway on arousal and escape behavior.
In further recordings (Figure 8A), activation of the insula–BLA pathway affected 42% of BLA neurons and reduced pupil size (Figures 8B–E); LFP power analysis showed no significant changes, contrasting with activation of the SC–MD pathway, suggesting that subcortical pathways play a more significant role than cortical pathways in regulating escape responses (Figures 8D–F).
Figure 8. Insula–BLA pathway modulates BLA oscillations.
Summary
This study found that rodents exhibit two behavioral response types to repeated visual threat stimuli: “persistent escape” (T1) and “rapid habituation” (T2). The T1 group showed larger pupil size, higher escape rates, and shorter response latency, whereas the T2 group exhibited the opposite characteristics along with enhanced non-selective attention. The SC and insular cortex project to the BLA via distinct pathways. The SC–VTA–BLA pathway mediates T1 behavior, while the SC–MD–BLA and insula–MD–BLA pathways mediate T2 behavior. The MD acts as a key hub integrating inputs to regulate arousal and defensive responses. This research provides important insights for understanding emotional processing, behavioral adaptability, and the treatment of related neuropsychiatric disorders.
The viral tools used in this study were provided by Brain Case.