Client Article | Cell Metabolism | Liu Danqian’s Team at the Institute of Neuroscience, CAS, Decodes the Oxidation-Sleep Code in Substantia Nigra Neurons
Release time:2025-09-24 13:42:32
The molecular mechanism underlying sleep need remains unclear. Oxidative stress has been proposed as a potential contributor to sleep regulation, but evidence in mammals is limited. In Drosophila, oxidative stress induces sleep via mitochondrial pathways in neurons. However, dynamic monitoring of reactive oxygen species (ROS) in the mammalian brain is challenging, and direct evidence has been lacking.
On May 15, 2025, Dr. Liu Danqian’s team at the Institute of Neuroscience, Center for Excellence in Brain Science and Intelligence Technology (CEBSIT), Chinese Academy of Sciences, published a study titled “Hydrogen peroxide in midbrain sleep neurons regulates sleep homeostasis” in Cell Metabolism. The study revealed a positive correlation between cytosolic hydrogen peroxide (H₂O₂) levels in midbrain sleep-active neurons of the substantia nigra and sleep pressure. Mild increases in H₂O₂ promote sleep initiation via activation of TRPM2 channels, whereas excessive H₂O₂ causes fragmented sleep and neuroinflammation.
This research was led by Ph.D. candidate Tian Yujing under the supervision of Dr. Liu Danqian at CEBSIT. Significant contributions were also made by Dr. Kang Luwei from CEBSIT, Dr. Deng Juan from the Institute of Brain Science, Fudan University, and Ngoc T. Ha.
Sleep Deprivation Leads to ROS Accumulation in Sleep-Promoting Brain Regions of Mice
Previous studies using brain homogenates or frozen sections failed to detect ROS accumulation following sleep deprivation. However, in situ analysis of the locus coeruleus (LC) revealed signs of oxidative stress. Traditional methods lack the high spatiotemporal resolution necessary for accurate in vivo ROS measurement, making it difficult to capture the real-time dynamics of specific brain regions.
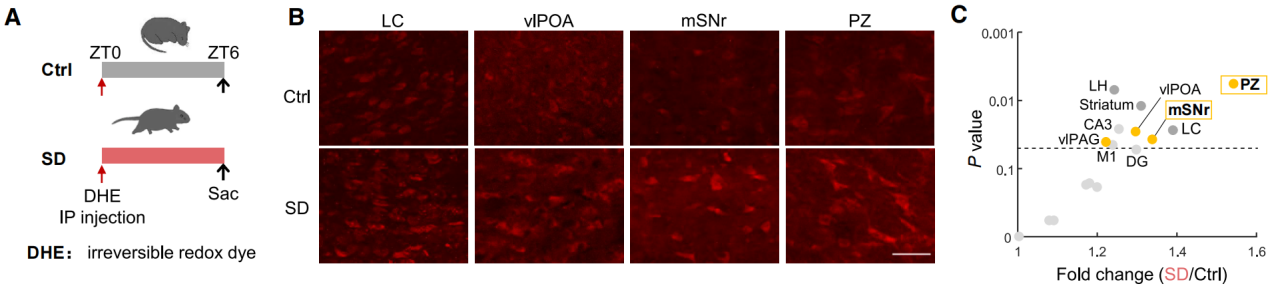
To address this challenge, the authors conducted whole-brain in situ detection of ROS in sleep-deprived mice. At Zeitgeber Time 0 (ZT0), mice received an intraperitoneal injection of dihydroethidium (DHE)—a superoxide-sensitive dye that crosses the blood-brain barrier and emits red fluorescence upon oxidation. Following injection, mice were either subjected to 6 hours of sleep deprivation (SD; ZT0–6) or left undisturbed (Figure 1A), after which perfusion and DHE analysis were performed.
Quantitative analysis of red fluorescence intensity across brain regions confirmed an increase in DHE signal in the locus coeruleus, consistent with prior findings (Figures 1A–1C). Notably, two key sleep-promoting regions—the parafacial zone (PZ) and the substantia nigra pars reticulata (SNr)—also showed significant ROS accumulation.
Figure 1. Sleep deprivation induces ROS accumulation in sleep-promoting brain regions of mice.
Cytosolic H₂O₂ in SNr Sleep Neurons Serves as an Indicator of Sleep Pressure During Prolonged Wakefulness
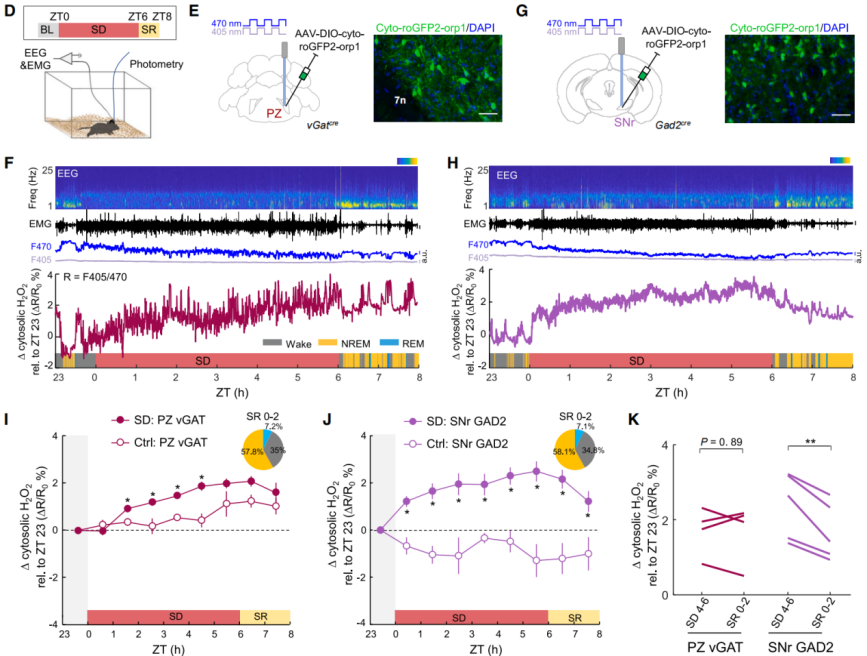
To investigate the relationship between H₂O₂ as a physiological signaling molecule and sleep (Figure 2D), AAV-DIO-cyto-roGFP2-Orp1 was injected into the parafacial zone (PZ) of vGAT-Cre mice or the SNr of Gad2-Cre mice (Figures 2E–2H). The roGFP2-Orp1 sensor enables the detection of physiological levels of H₂O₂ in brain tissue. PZ vGAT and SNr GAD2 neurons are sleep-related neurons. Using a fiber-optic system to rapidly alternate between 405 nm and 470 nm excitation wavelengths, the F405/F470 fluorescence ratio reflects fluctuations in H₂O₂ levels.
The results showed that during SD, cytosolic H₂O₂ in SNr neurons rapidly increased and reached a plateau at approximately 2.4%, consistent with the saturation curve of sleep pressure (Figure 2J). After sleep recovery (SR), H₂O₂ levels in SNr significantly decreased, while those in PZ showed no obvious changes, suggesting a reversible correlation between SNr H₂O₂ accumulation and sleep pressure (Figures 2I–2K). In contrast, H₂O₂ in the PZ gradually increased during the later phase of SD but remained unchanged during recovery, indicating that the accumulated ROS may include other species such as superoxide rather than pure H₂O₂ (Figure 2I). In normal control mice, H₂O₂ in the SNr decreased during the light phase (resting phase) and increased during the dark phase (active phase), indicating regulation by circadian rhythms (Figure 2J).
Figure 2. Cytosolic H₂O₂ in SNr sleep neurons serves as an indicator of sleep pressure during prolonged wakefulness.
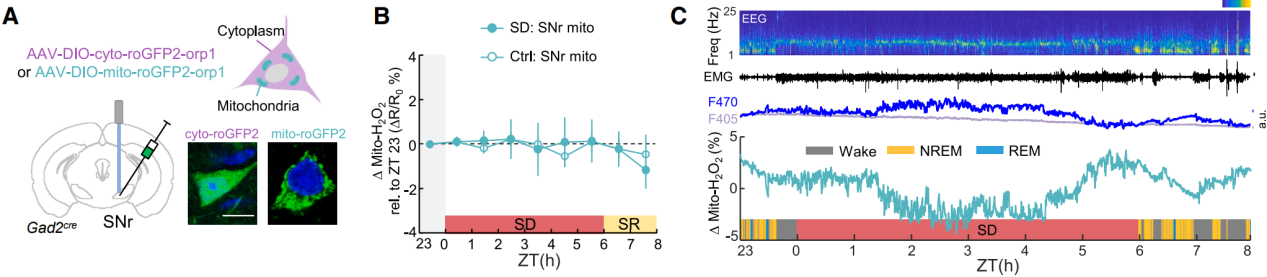
A mitochondria-targeted sensor (mito-roGFP2-Orp1) revealed that mitochondrial H₂O₂ levels did not show sustained changes during SD (Figure 3A), ruling out mitochondria as the source of cytosolic H₂O₂ (Figures 3B–3C). Inhibition of NADPH oxidase (NOX) blocked the accumulation of cytosolic H₂O₂, indicating that NOX is the primary source of H₂O₂ production in SNr neurons.
In conclusion, cytosolic H₂O₂ accumulation in SNr sleep neurons is a reliable marker of sleep pressure during prolonged wakefulness.
Figure 3. Mitochondrial H₂O₂ does not contribute to cytosolic H₂O₂ accumulation during SD.
Cytosolic H₂O₂ in SNr Sleep Neurons Tracks Spontaneous Wakefulness
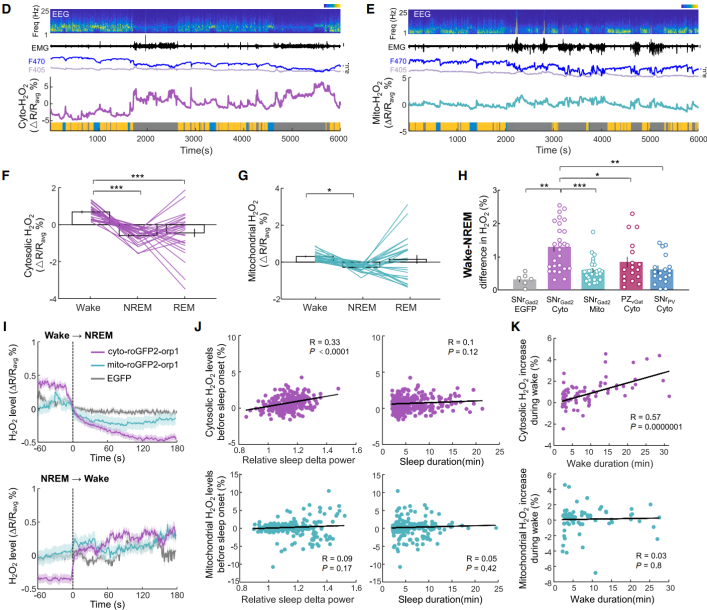
Next, the study examined whether H₂O₂ could track spontaneous sleep–wake cycles under undisturbed conditions, and aimed to distinguish functional differences between cytosolic and mitochondrial H₂O₂. The dynamics of both cytosolic and mitochondrial H₂O₂ in SNr GAD2 neurons of undisturbed mice were measured. During wakefulness, levels of both cytosolic and mitochondrial H₂O₂ were higher than during non-rapid eye movement (NREM) sleep, and both showed large fluctuations during rapid eye movement (REM) sleep (Figures 4D–4G). The difference in cytosolic H₂O₂ between wakefulness and NREM sleep was significantly greater (Figure 4H), suggesting that cytosolic H₂O₂ more sensitively reflects transitions between wakefulness and sleep. Following sleep onset, both mitochondrial and cytosolic H₂O₂ levels steadily declined, but only cytosolic H₂O₂ at sleep initiation correlated with sleep δ power (Figures 4I–4J). Upon awakening, mitochondrial H₂O₂ showed almost no increase, whereas cytosolic H₂O₂ gradually rose with increasing wake duration and showed a positive correlation with total wake time (Figures 4I and 4K). This correlation was not observed in PZ sleep neurons or other SNr neuronal populations (Figure 4H). These findings indicate that cytosolic H₂O₂ in SNr sleep neurons also reflects the dynamics of spontaneous sleep pressure, tracks wake duration, and influences sleep intensity.
H₂O₂ Is a Key Signaling Molecule in Sleep Regulation
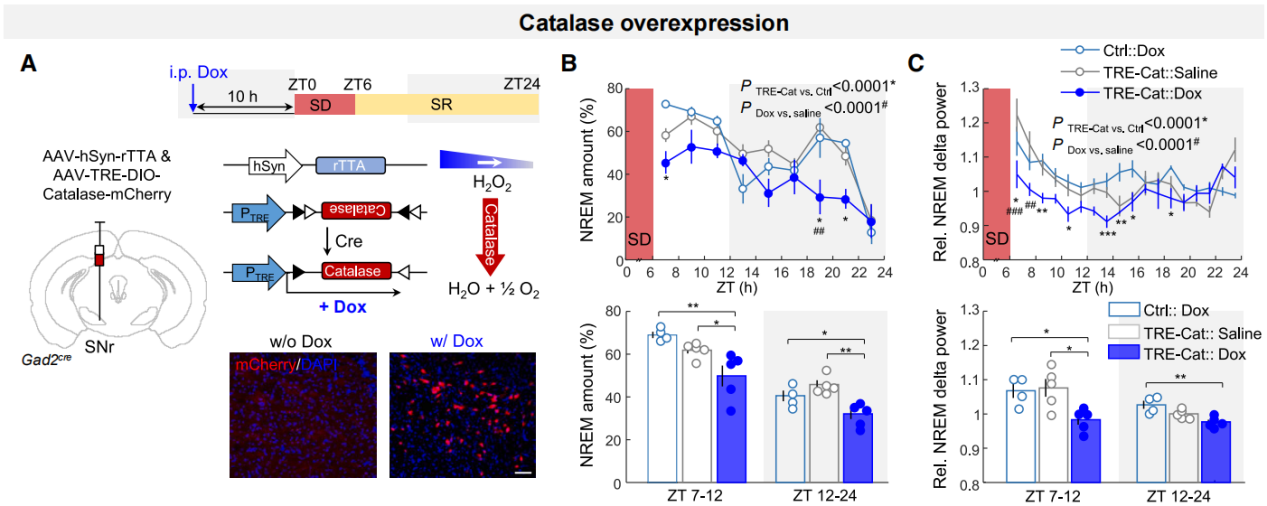
To determine whether H₂O₂ actively regulates sleep, a tetracycline-inducible system was used to specifically overexpress catalase (an enzyme that degrades intracellular H₂O₂) in SNr GAD2 neurons (Figure 5A). Doxycycline (dox) was administered more than 12 hours before sleep recording to ensure sufficient expression of catalase. Compared to control conditions, dox-induced catalase overexpression significantly reduced compensatory sleep during the 18-hour SR period and decreased immediate post-SD sleep δ power (Figures 5B–5C). These findings suggest that cytosolic H₂O₂ in SNr sleep neurons is essential for regulating both the quantity and intensity of sleep, particularly during rebound sleep, underscoring its critical role in maintaining sleep homeostasis.
Figure 5. H₂O₂ is a key signaling molecule in sleep regulation.
Tuning H₂O₂ Production in SNr Sleep Neurons Promotes Sleep Initiation
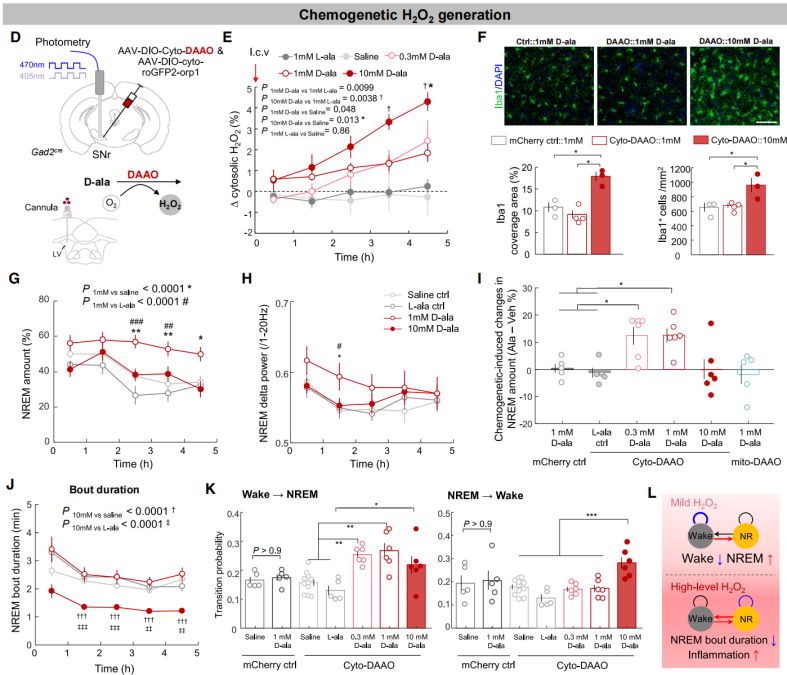
To determine whether cytosolic H₂O₂ directly promotes sleep, AAV-DIO-cyto-DAAO (DAAO, an enzyme that produces H₂O₂ in response to D-alanine [D-ala]) and AAV-DIO-cyto-roGFP2-Orp1 were co-injected into the SNr of Gad2-Cre mice to simultaneously induce and monitor H₂O₂ production (Figure 6D). The study found that intracerebroventricular (i.c.v.) infusion of D-ala in DAAO-expressing mice induced dose-dependent increases in H₂O₂: 0.3–1 mM D-ala caused a mild elevation, while 10 mM D-ala led to a more pronounced rise (Figures 6D–6E). Immunostaining for the microglial activation and inflammation marker Iba1 showed increased signal following 10 mM D-ala infusion, but not after 1 mM, indicating that low-dose D-ala induced mild H₂O₂ elevation without notable inflammation, whereas high-dose D-ala triggered excessive H₂O₂ and neuroinflammation (Figure 6F).
The authors then investigated how elevated H₂O₂ affects sleep. They found that a mild increase in H₂O₂ within SNr sleep neurons significantly enhanced NREM sleep (Figures 6G–6J), primarily by facilitating sleep initiation (Figure 6K). Moreover, infusion of 1 mM D-ala in mice expressing mito-DAAO in the SNr or cyto-DAAO in the PZ had no effect on sleep, confirming the specific role of cytosolic H₂O₂ in SNr sleep neurons (Figure 6I). In contrast, high levels of H₂O₂ in the SNr (via 10 mM D-ala) did not change total sleep time (Figures 6G–6I) but led to sleep fragmentation, as evidenced by shorter NREM bouts and increased NREM-to-wake transitions (Figures 6J–6K). This effect may underlie sleep disturbances observed in aging and neurodegenerative diseases linked to brain oxidative stress and inflammation.
In summary, cytosolic H₂O₂ in SNr sleep neurons plays a dual role: at mild levels, it promotes sleep initiation, while at excessive levels, it induces sleep fragmentation (Figure 6L).
Figure 6. Tuning H₂O₂ production in SNr sleep neurons promotes sleep initiation.
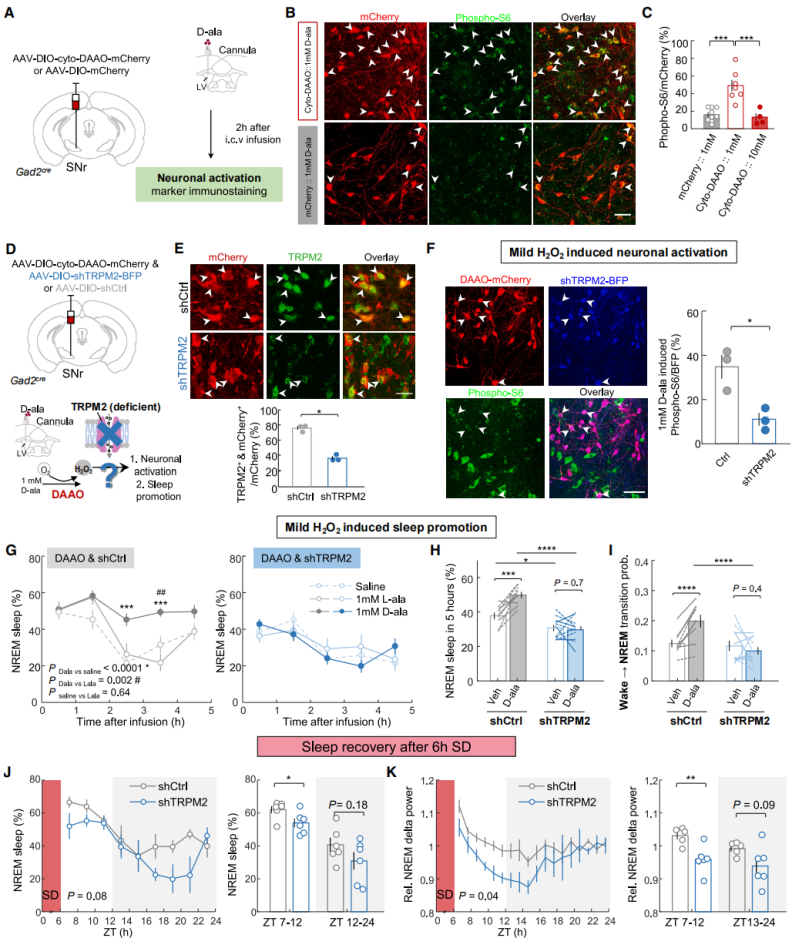
The neuronal activation and sleep-promoting effects induced by mild H₂O₂ in sleep-active substantia nigra neurons depend on TRPM2
Next, the mechanism by which cytosolic H₂O₂ regulates sleep initiation was investigated. Immunostaining for the neuronal activation marker phospho-S6 revealed that administration of 1 mM D-alanine (D-ala) significantly increased phospho-S6-positive signals in DAAO-expressing neurons (Figures 7A–7C), indicating that mild H₂O₂ enhances neuronal activity. Electrophysiological recordings further showed that DAAO-mediated H₂O₂ production reversibly increased neuronal excitability. Previous studies have shown that H₂O₂ can enhance neuronal excitability by activating transient receptor potential (TRP) channels. The involvement of H₂O₂-sensitive TRP channels was then examined. It was found that TRP melastatin 2 (TRPM2) had the highest mRNA expression levels in the SNr, and 83.7% of GAD2 neurons expressed this channel. AAV-DIO-cyto-DAAO and AAV-DIO-shTRPM2 were co-injected into the SNr to perform gene knockdown (Figures 4D–4F). Results showed that in scrambled control mice, 1 mM D-ala promoted NREM sleep (Figures 7G–7I), but this sleep-promoting effect and neuronal excitability enhancement were completely abolished in shTRPM2 mice (Figures 7F–7I). TRPM2 knockdown also reduced sleep amount and δ power during sleep recovery (Figures 7J–7K), and decreased baseline NREM sleep during the light phase.
In summary, H₂O₂ dynamics in sleep-active substantia nigra neurons can track sleep pressure and regulate sleep homeostasis through a TRPM2-dependent mechanism.
Figure 7. The effects of mild H₂O₂ elevation on neuronal activation and sleep promotion depend on TRPM2
Summary
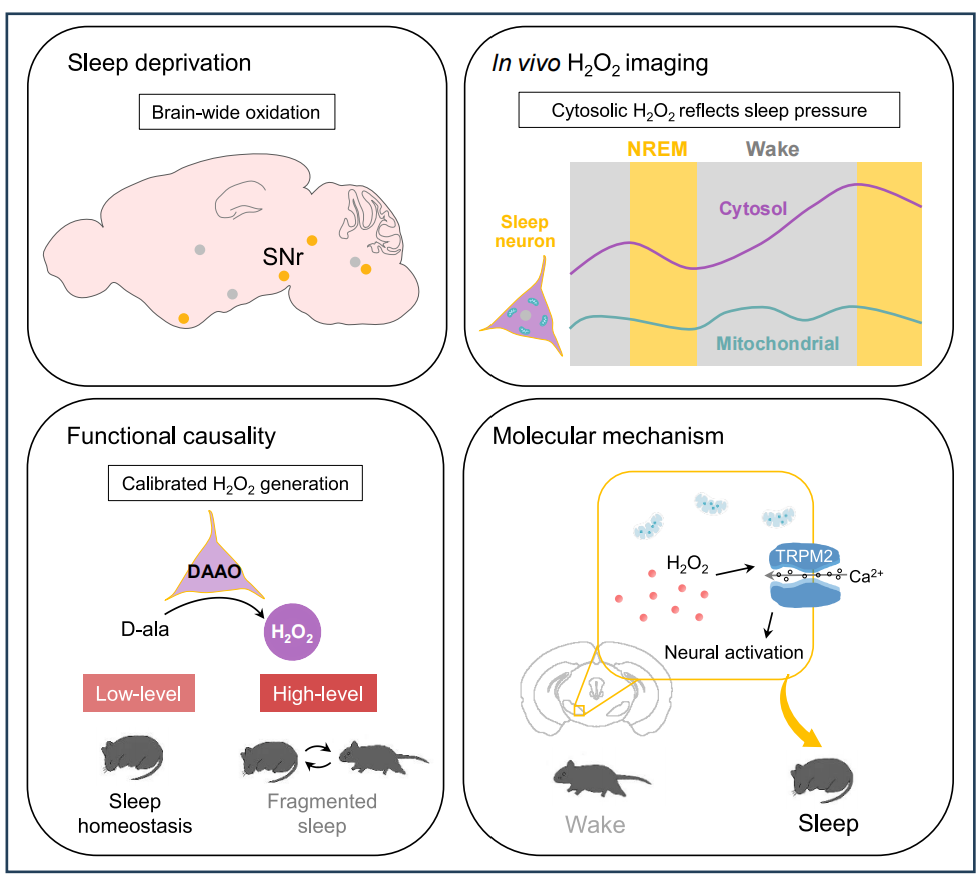
This study is the first to reveal the dual role of cytosolic H₂O₂ as an “oxidative homeostasis signal”: at physiological levels, it transforms oxidative stress into a sleep-promoting drive, whereas pathological overproduction disrupts sleep architecture. These findings offer new targets for understanding sleep regulation mechanisms and oxidative stress-related sleep disorders, such as aging and neurodegenerative diseases.
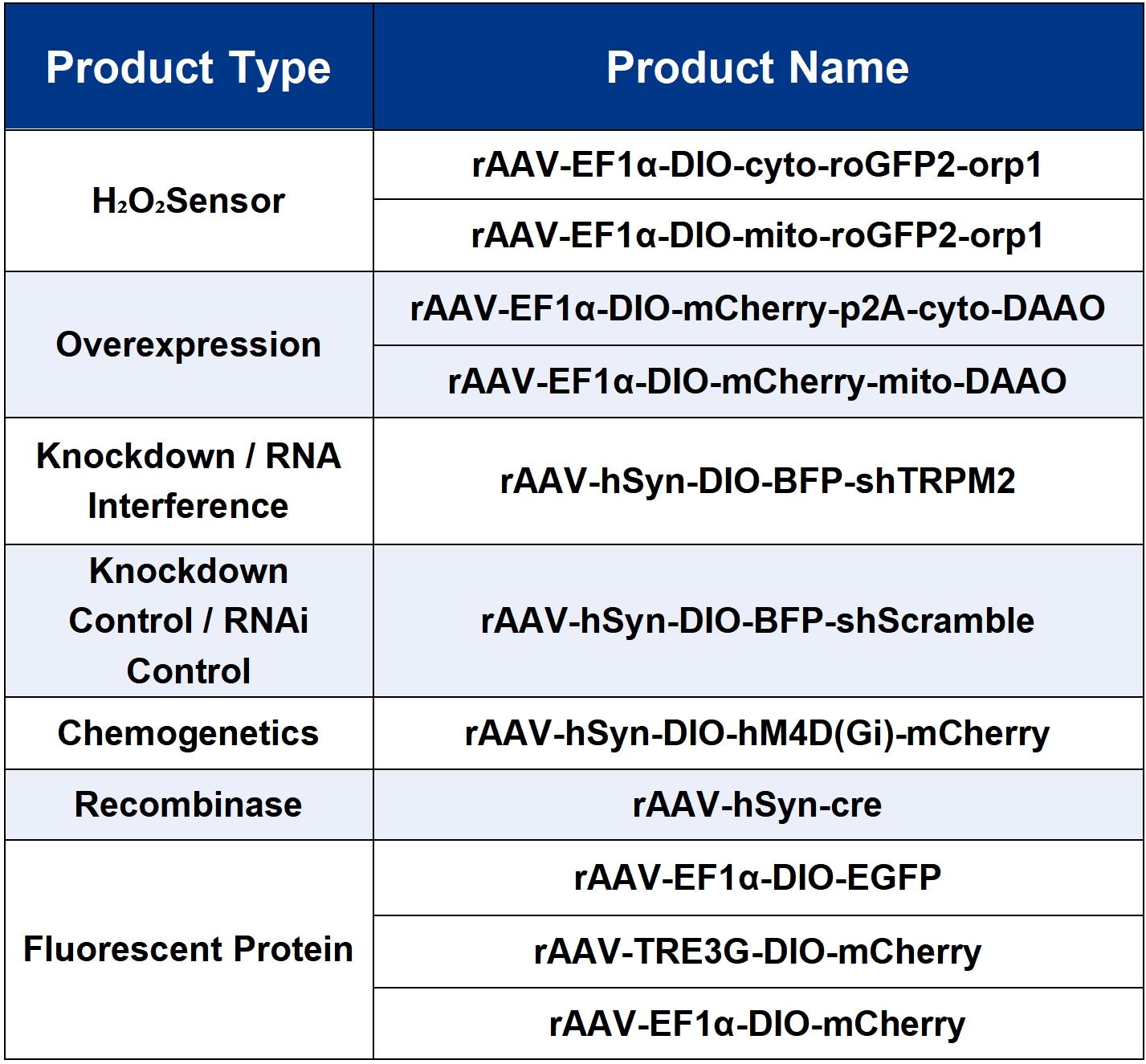
The viral tools used in this study are all available from Brain Case Biotech.
Brain Casecan provide customers with a full range of vector construction, virus packaging services. If you are interested in customized services, please contact bd@ebraincase.com for details or to place an order.
Service Type :
Select the service you'd like to purchase.
Order Information(Premade-AAVs)
Please provide us some information about the service you'd like to order.
Order Information(Custom AAV/Lentivirus)
Please provide us some information about the service you'd like to order.
Order Information(Others)
Please provide us some information about the service you'd like to order.