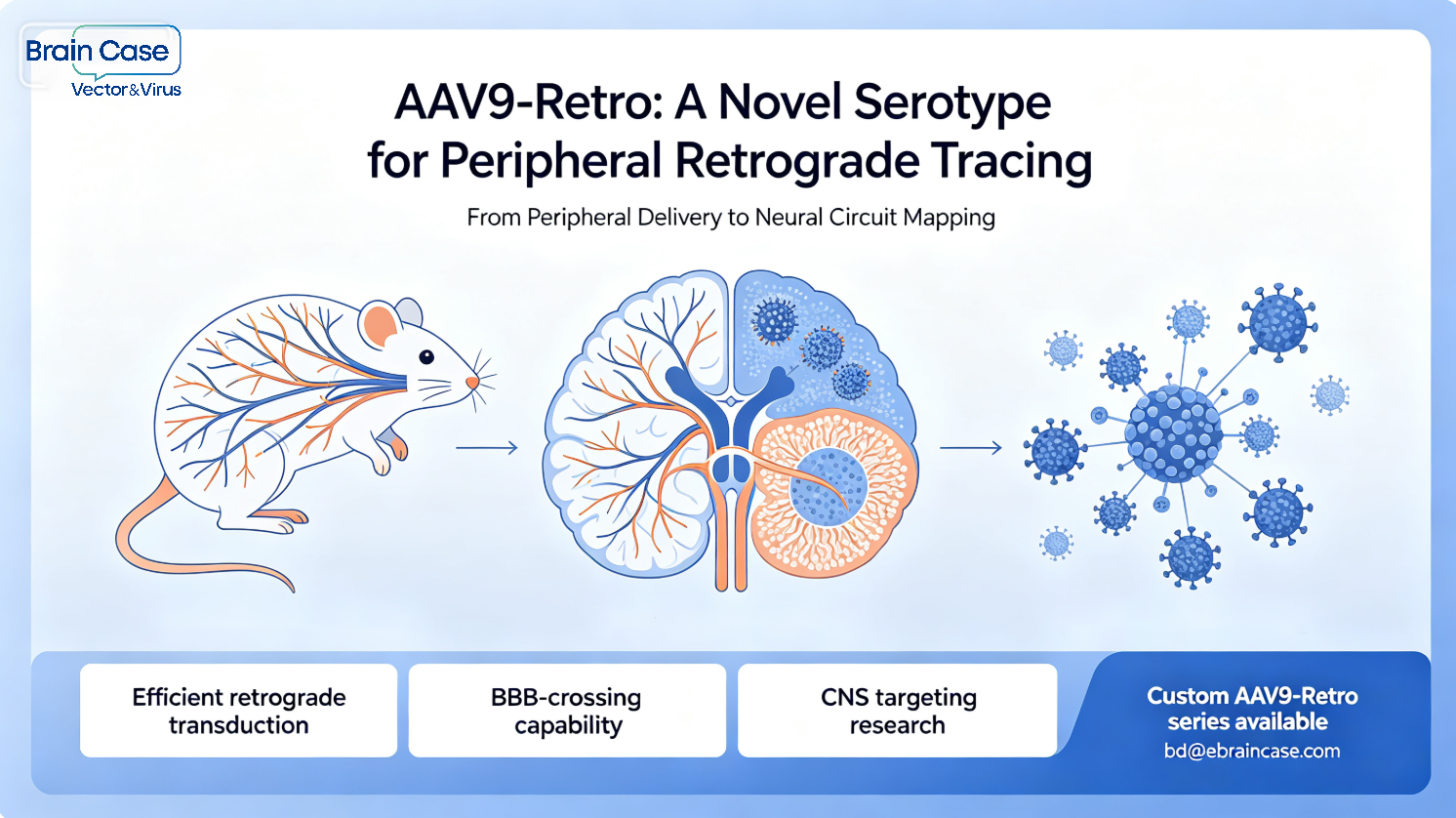
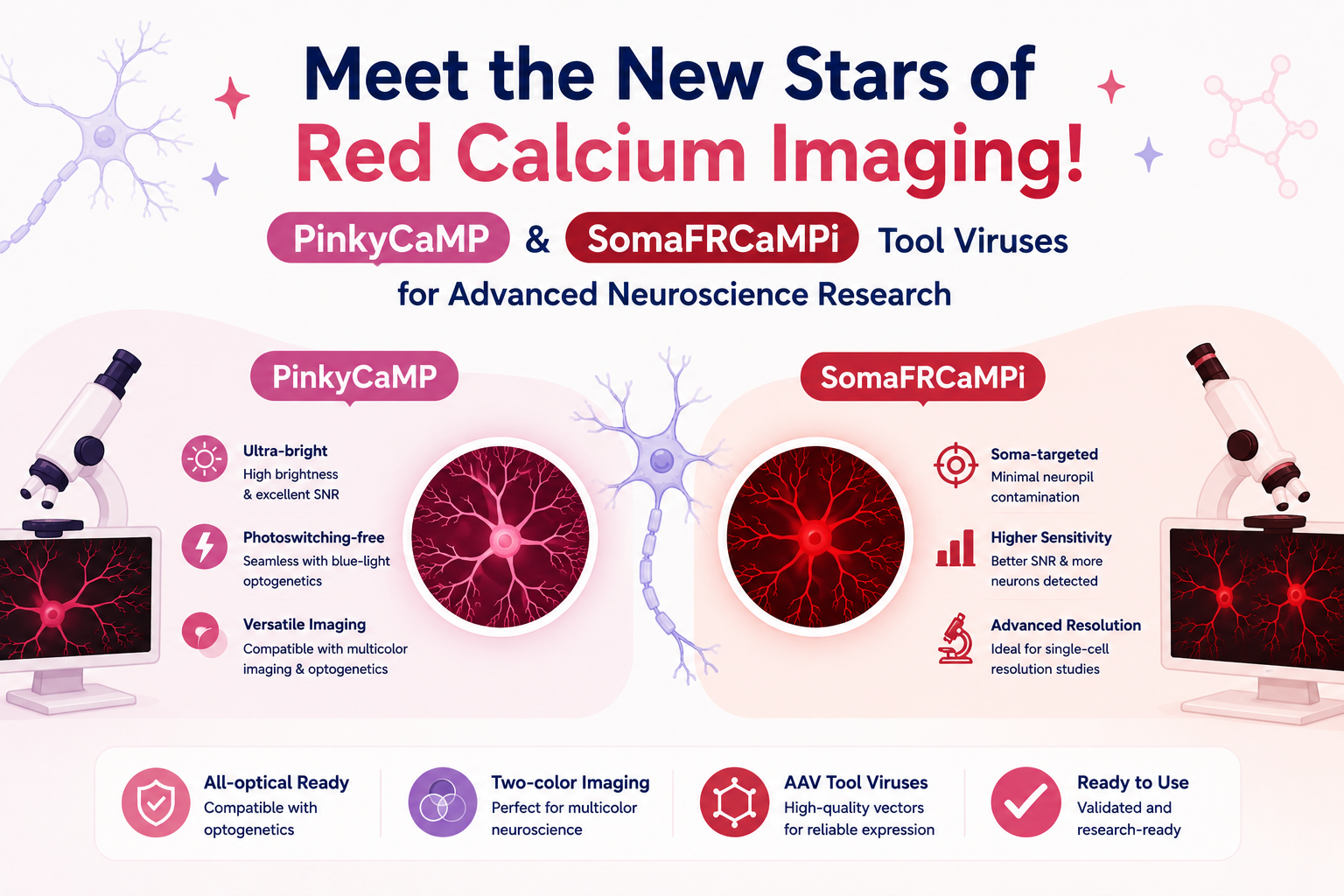
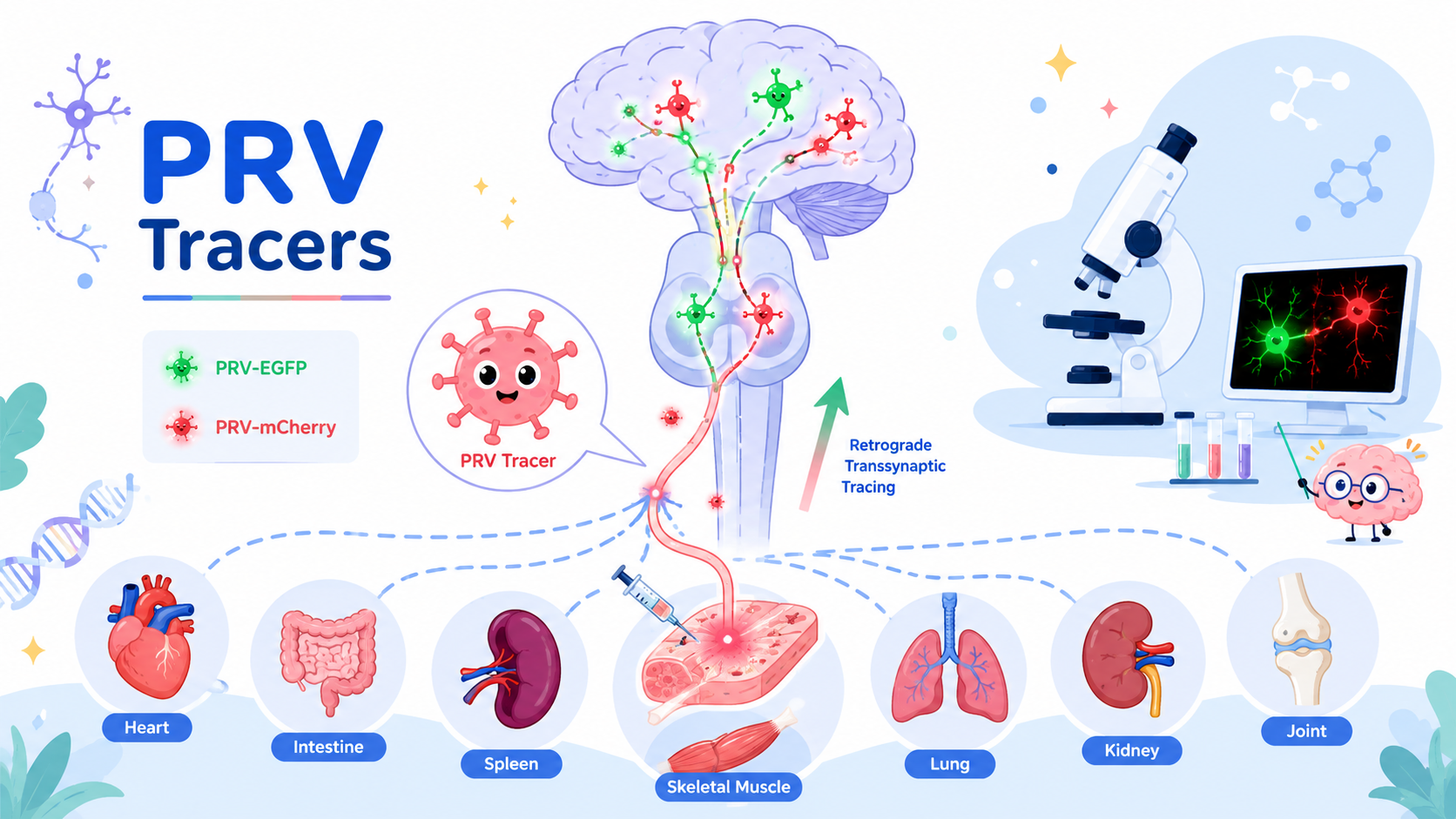
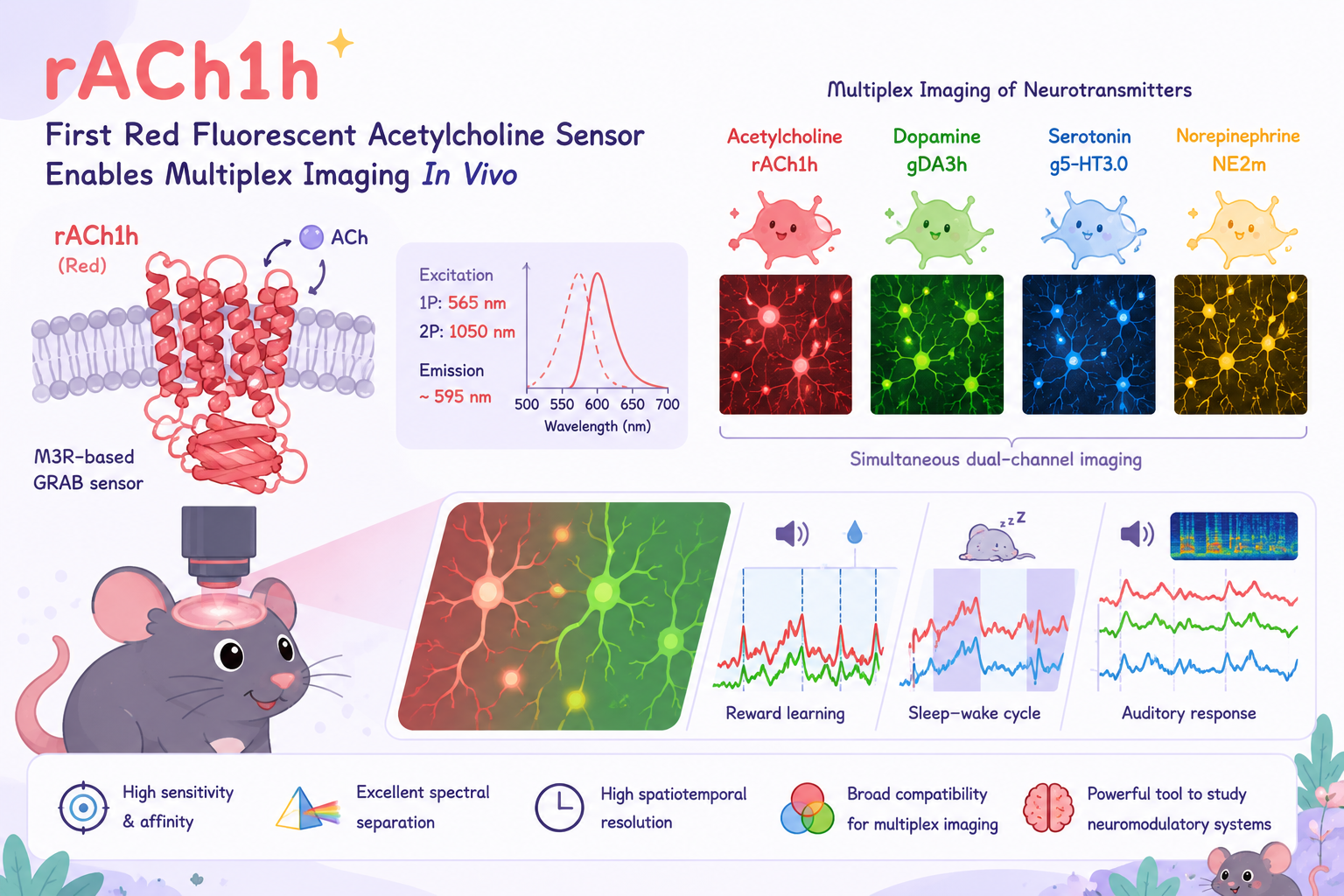
GABAergic neurotransmission plays a critical role in shaping neural circuit dynamics and maintaining the balance between excitation and inhibition in the brain. Abnormal GABA signaling is associated with various neurological and psychiatric disorders, including epilepsy, schizophrenia, and autism.
However, current techniques for measuring GABA levels face limitations in sensitivity and spatiotemporal resolution. For example, microdialysis offers low temporal resolution, limited to the scale of minutes, while electrophysiological methods are not suitable for monitoring large populations of neurons and struggle to distinguish GABAergic signals from other hyperpolarizing currents.
Previously developed genetically encoded fluorescent sensors, such as iGABASnFR1, enabled monitoring of GABAergic activity but had limited sensitivity and dynamic range. In cultured neurons, iGABASnFR1 had a half-maximal effective concentration (EC50) of approximately 30 μM—where EC50 refers to the concentration at which the sensor elicits 50% of its maximal response, with lower EC50 indicating higher affinity for GABA. Its maximal fluorescence change (ΔF/F) was around 0.6, making its overall performance inadequate for high-resolution, photon-limited imaging applications.
Research Findings
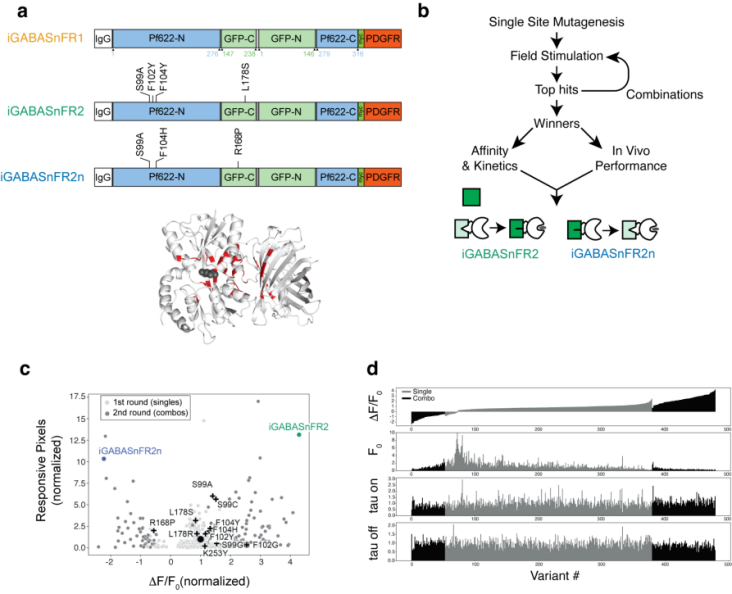
Building upon iGABASnFR1, researchers performed near-saturation mutagenesis on 39 amino acid residues located near the GABA binding site, the protein hinge region, cpSFGFP, and the interface between the ligand-binding domain and cpGFP. By combining beneficial single-point mutations to improve sensor performance and conducting two rounds of screening, they identified an optimal variant with enhanced dynamic range and expression: iGABASnFR2 (increased fluorescence upon GABA binding), as well as a reverse-signaling variant, iGABASnFR2n (decreased fluorescence upon GABA binding) (Figure 1).
Figure 1. Field stimulation screening used to improve iGABASnFR.
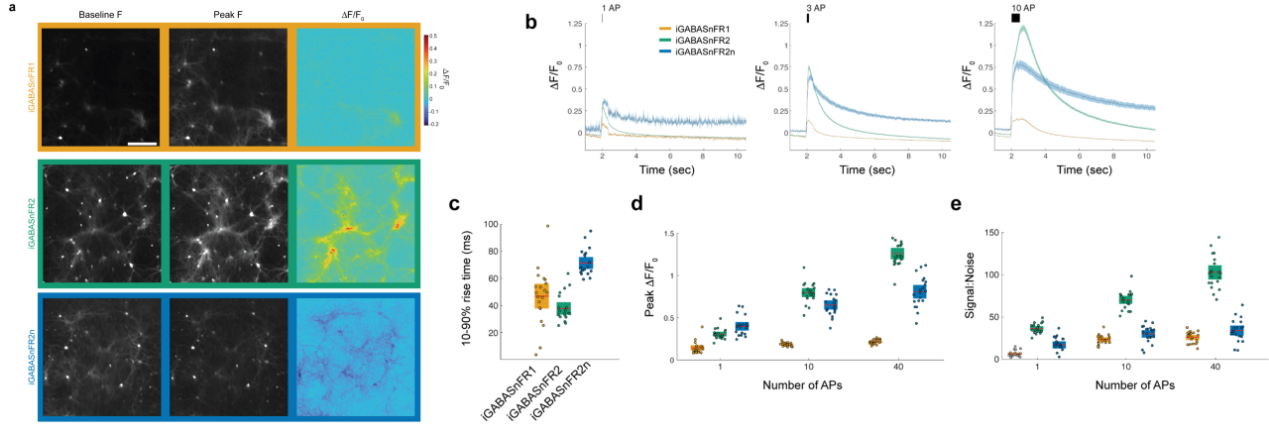
In terms of response kinetics, peak ΔF/F, and signal-to-noise ratio (SNR), iGABASnFR2 significantly outperformed iGABASnFR1 across all field stimulation conditions. When stimulated with 10 action potentials (APs), iGABASnFR2 achieved a peak ΔF/F of 0.80 ± 0.12, which is 4.2 times higher than that of iGABASnFR1, and an SNR of 70.5 ± 10.7, three times greater than iGABASnFR1. Its rise time was 38 ± 10 ms, indicating faster kinetics. The reverse-signal variant iGABASnFR2n had a peak ΔF/F 3.2 times higher than iGABASnFR1, with an SNR 30% higher (Figure 2). These results suggest that both iGABASnFR2 and iGABASnFR2n demonstrate broadly improved performance in detecting GABA dynamics during synaptic transmission.
Figure 2. Characterization of iGABASnFR variants in cultured neurons.
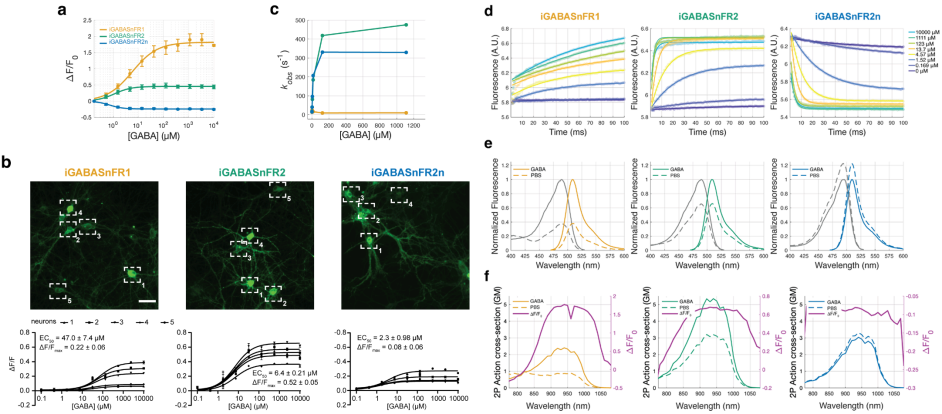
For in vivo applications, the effectiveness of a sensor depends on its affinity for GABA and its binding kinetics. In terms of affinity, iGABASnFR2 showed a cellular EC50 of 6.4 ± 0.21 μM, reflecting a 7-fold higher affinity than iGABASnFR1, significantly enhancing its GABA-binding capability. Kinetically, iGABASnFR1 exhibited biphasic behavior, while iGABASnFR2 and iGABASnFR2n followed single-exponential kinetics, indicating a simpler relationship between ligand binding and fluorescence change. Spectrally, the 1-photon and 2-photon excitation spectra of iGABASnFR2 and iGABASnFR2n were similar to that of iGABASnFR1, as were their apparent pKa values (Figure 3).
Figure 3. Biophysical properties of iGABASnFR variants.
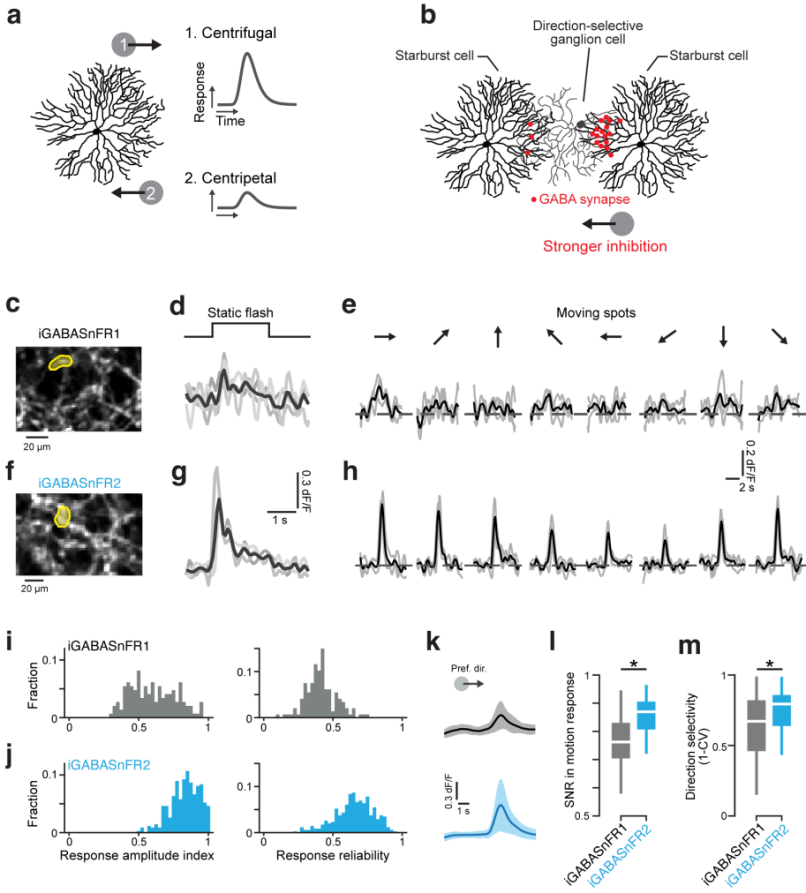
In studies of synaptic transmission in the retina, researchers used a Cre-Lox system to transduce starburst amacrine cells (SACs) with iGABASnFR1 or iGABASnFR2 via viral delivery. After 6–8 weeks, retinal tissues were dissected for two-photon imaging. While iGABASnFR1 showed only weak responses to full-field stimuli and struggled to detect single-trial responses or direction-selective GABA release, iGABASnFR2 enabled detection of responses to static flashes in single trials and clearly revealed direction-selective GABA release in response to motion stimuli. By calculating the Response Amplitude Index (RAI) and response reliability, researchers found that iGABASnFR2 significantly outperformed iGABASnFR1. The average response reliability of iGABASnFR2 was 0.66 ± 0.14, compared to 0.41 ± 0.11 for iGABASnFR1 (Figure 4). Moreover, iGABASnFR2 exhibited a much higher SNR and more accurate direction-selectivity measurements, supporting the hypothesis that SACs release GABA in a direction-selective manner in the retina.
Figure 4. iGABASnFR2 reports direction-selective GABA release in the retina.
Research Conclusion
As powerful tools for studying GABAergic inhibition in the nervous system, these sensors hold great promise for advancing our understanding of the mechanisms underlying neurological and psychiatric disorders associated with GABA signaling.
iGABASnFR2 and iGABASnFR2n exhibit significantly enhanced affinity for GABA and greatly improved detection sensitivity. In retinal experiments, iGABASnFR2 demonstrated outstanding performance, enabling the detection of more responses and providing more precise measurements of GABA release triggered by visual motion. This advancement offers crucial insights into the origin of direction selectivity in the retina.
Ⅱ. iGluSnFR4f/4s Glutamate Sensors: A Spatiotemporally Customizable Tool for Monitoring Synaptic Glutamate Dynamics
Research Background
Glutamate is the primary neurotransmitter in the central nervous system of vertebrates. Due to its low molecular abundance and brief synaptic residence time, optically measuring synaptic glutamate release—especially across multiple synapses in vivo—poses significant challenges. Although fluorescent glutamate indicators such as iGluSnFR can detect single synaptic vesicle release events, iGluSnFR3 lacks sufficient SNR for monitoring dozens of synapses simultaneously in vivo. As the number of imaged synapses increases, the total excitation power must be distributed among them, reducing SNR. Moreover, the voxel rate of microscopy (the number of voxels captured or processed per unit time) imposes a trade-off between sampling rate and the number of monitored synapses. These limitations highlight the need for brighter and more sensitive indicators to meet the growing demands of synaptic imaging.
Research Findings
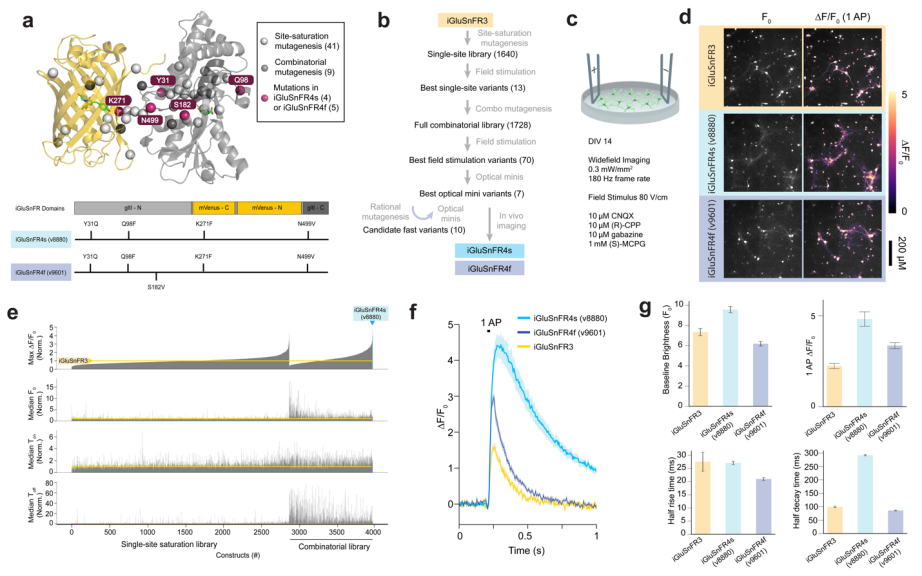
The researchers performed saturation mutagenesis at 41 previously identified sites in two iGluSnFR3 variants (iGluSnFR3.v857 and iGluSnFR3.v867), generating a library of 1,640 variants. These were screened in cultured neurons using field stimulation by measuring parameters such as baseline fluorescence intensity (F₀), peak response amplitude (∆F/F₀), and rise and decay times (Ton and Toff). From this, 12 promising point mutations were selected to construct a combination library of 1,728 variants. A generalized linear model (GLM) was used to analyze the individual and interactive effects of each mutation.
Subsequently, 70 variants were selected from the combination screening and tested for spontaneous synaptic glutamate release in TTX-silenced cultured neurons, identifying variants with rapid decay kinetics and improved signal-to-noise ratios. Finally, seven top candidates were evaluated in vivo in the mouse visual cortex. Using two-photon imaging to assess multiple performance metrics, two best-performing variants were identified: v8880 and v9601, named iGluSnFR4s (slow deactivation) and iGluSnFR4f (fast deactivation), respectively (Fig. 5).
Figure 5 Screening of iGluSnFR variants in primary neuronal cultures.
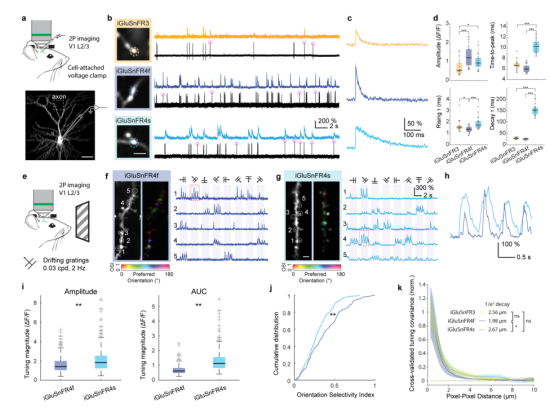
In the mouse visual cortex, sparse expression of iGluSnFR variants in primary visual cortex (V1) neurons was achieved via single-cell electroporation. Whole-cell patch-clamp recordings and two-photon imaging were used to characterize glutamate transients triggered by single action potentials (APs) in the recorded neurons. Both iGluSnFR4f and iGluSnFR4s effectively detected glutamate release events induced by single APs, demonstrating higher sensitivity and faster decay kinetics. Additionally, the two probes showed clear distinctions in signal decay characteristics: iGluSnFR4f was better suited for tracking rapid synaptic dynamics, while iGluSnFR4s offered a unique advantage in prolonged signal duration (Fig. 6a–d).
To evaluate spatial specificity, the researchers presented mice with drifting grating visual stimuli and imaged dendritic responses in V1 neurons. Both iGluSnFR4f and iGluSnFR4s reported single-synapse signals with high signal-to-noise ratio and specificity, with signals localized to dendritic spines. iGluSnFR4f exhibited sharper temporal responses, while iGluSnFR4s produced larger peak amplitudes and integrated response areas (Fig. 6e–k).
Figure 6 Characterization of iGluSnFR4 in the mouse visual cortex.
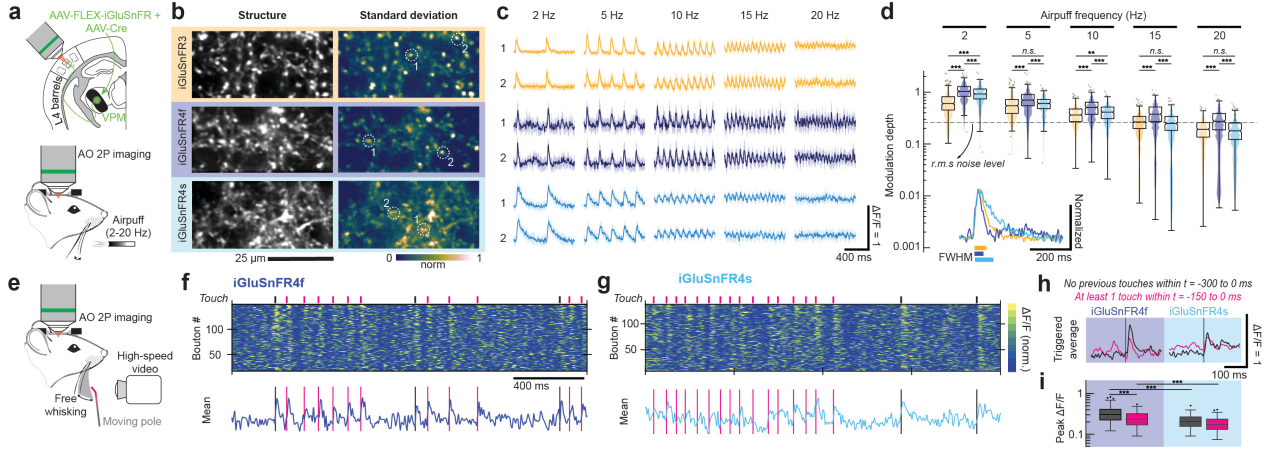
In the ventral posteromedial thalamus (VPM), iGluSnFR variants were injected and imaged in thalamocortical axons within the mouse primary somatosensory cortex (vS1) using an adaptive optics two-photon (AO-2P) microscope. During rhythmic air-puff stimulation and voluntary whisker movement tasks, both variants elicited rapid-onset responses across stimulation frequencies ranging from 2 to 30 Hz. Notably, iGluSnFR4f maintained detection capability up to 20 Hz. In the freely whisking condition, iGluSnFR4s—due to its slower decay—underperformed in capturing fast, repeated touch events. In contrast, iGluSnFR4f produced distinguishable responses to different touch patterns (e.g., no touch in the previous 300 ms vs. at least one touch in the previous 150 ms) with higher average amplitude, making it more suitable for monitoring sensory input in the rodent somatosensory cortex.
Figure 7 Imaging fast dynamics in thalamocortical axons using AO-2P microscopy.
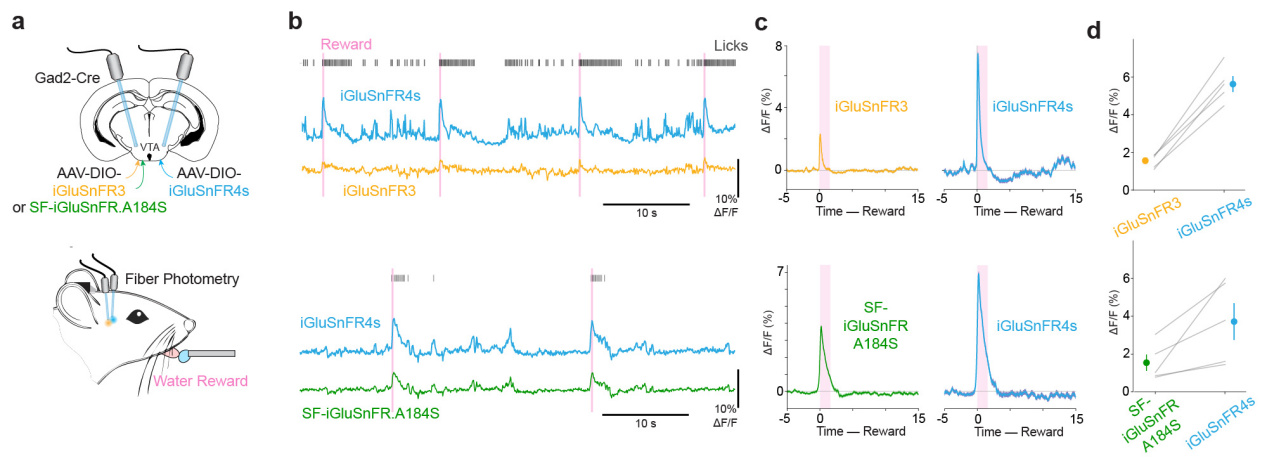
In the ventral tegmental area (VTA) of the midbrain, iGluSnFR4s and control probes were expressed in GABAergic neurons. Fiber photometry was used to record responses to water reward delivery. iGluSnFR4s produced significantly stronger responses than both iGluSnFR3 and SF-iGluSnFR, with amplitudes of 5.62±0.42 vs. 1.56±0.16 and 3.72±0.92 vs. 1.54±0.44, respectively—demonstrating superior signal detection capability in deep brain regions.
Figure 8 Fiber photometry in deep brain regions using iGluSnFR4s.
Research Conclusion
iGluSnFR4f and iGluSnFR4s demonstrate enhanced sensitivity and tunable inactivation kinetics. iGluSnFR4f excels in tracking rapid synaptic dynamics, while iGluSnFR4s shows distinct advantages in applications requiring high signal amplitude, such as two-photon imaging and fiber photometry. Both probes are capable of detecting a variety of natural synaptic transmission events, making them powerful tools for investigating neuronal computation, synaptic physiology, and the underlying mechanisms of brain disorders. Their development significantly advances research in the field of neuroscience.
Brain Case Glutamate and GABA Sensor Product List: