Literature Review | Comprehensive Summary of 4 Major Photosensitive Receptor Tools! Full Landscape of OptoXR Technology: Precise Control of Neural Circuits and Behavior via D1R/MOR/CB1R/A2AR
Release time:2025-09-25 17:14:37
Photosensitive Dopamine Receptor 1 (opto-D1)
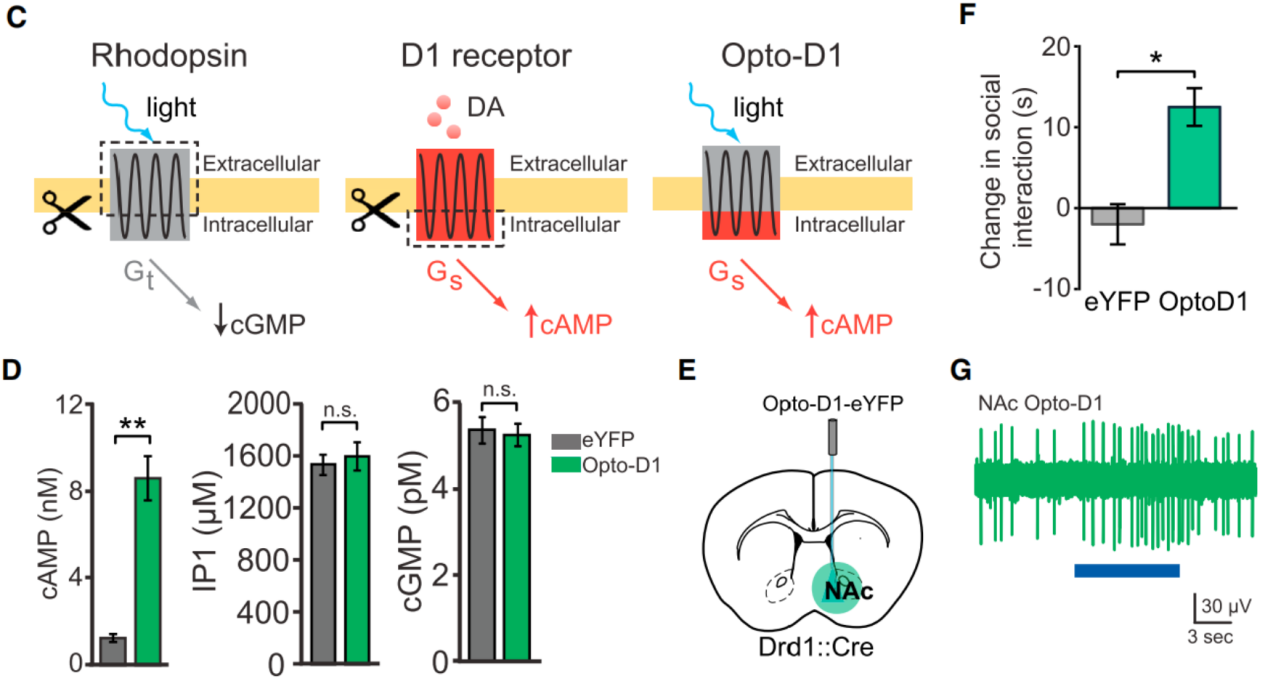
On July 19, 2014, researchers from the Howard Hughes Medical Institute published a study in Cell titled “Natural Neural Projection Dynamics Underlying Social Behavior”.Using fiber photometry to record neural activity in freely moving mice, combined with optogenetics, they discovered that dopaminergic projections from the ventral tegmental area (VTA) to the nucleus accumbens (NAc) encode and predict social interactions.Optogenetic manipulation of this projection bidirectionally regulated social behavior, and the effect depended on D1 receptors (D1R) in the NAc. Further, by employing OptoXR technology, the team engineered a chimeric Opto-D1 receptor and directly stimulated D1R-positive medium spiny neurons (MSNs) in the NAc, confirming that D1R-mediated signaling is sufficient to drive social behavior. These findings reveal the circuit-level dynamics underlying social behavior. To test the sufficiency of D1R-expressing MSNs in social behavior, the researchers constructed a light-activated chimeric Opto-D1 receptor (replacing the intracellular loops of rhodopsin with D1R sequences, Fig. 1C).In HEK cells expressing Opto-D1-eYFP, light selectively upregulated the cAMP (Gs) signaling pathway without affecting IP3 (Gq) or cGMP (Gt) pathways (Fig. 1D). By injecting a Cre-dependent Opto-D1 adeno-associated virus into the NAc of Drd1::Cre mice, expression was restricted to D1R MSNs (Fig. 1E).Results showed that 473 nm light significantly increased social interaction (Fig. 8F) without affecting locomotion or novel object exploration.Optrode recordings revealed that light increased NAc multi-unit activity by ~3-fold (Fig. 1G), matching the firing changes observed during natural social interaction or VTA optogenetic activation.
Figure 1. Role of postsynaptic NAc D1 neurons and receptors in natural and VTA-driven social behavior.
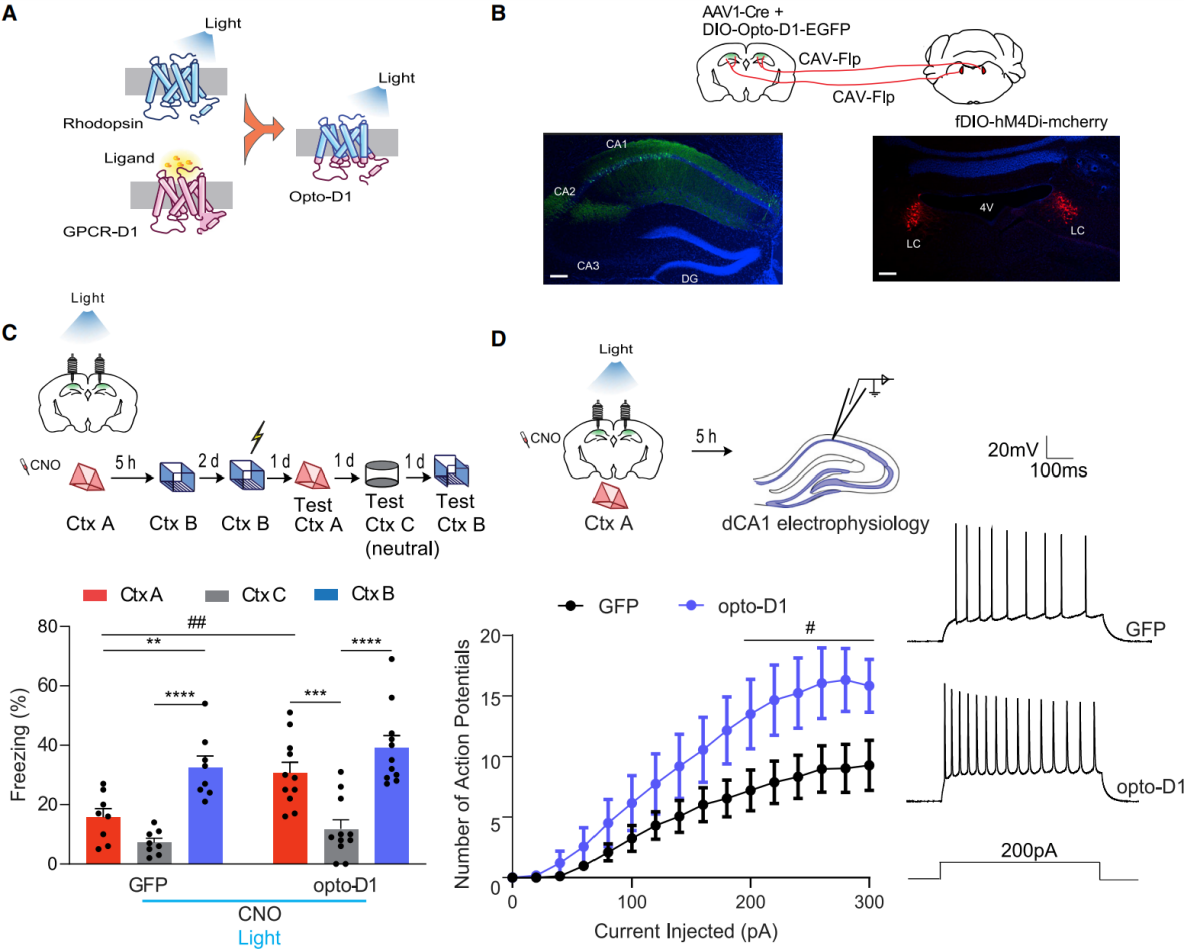
On October 19, 2022, researchers from the University of California published a study in Neuron titled “A locus coeruleus–dorsal CA1 dopaminergic circuit modulates memory linking”.They identified a dopaminergic pathway from the locus coeruleus (LC) to dorsal CA1 (dCA1) that specifically regulates the linking of temporally close contextual memories without affecting memory formation itself.Using opto-D1 to activate D1 receptor signaling in dCA1 neurons (Fig. 2A–B), they inhibited LC neurons projecting to dCA1 with CNO (administered 30 min before exploration of Context A). During Context A exploration, blue light (473 nm, 50 s on/10 s off, 10 mW) was applied to activate D1 receptors in dCA1, after which mice were returned to their home cage. Five hours later, the mice explored Context B with no CNO or light (Fig. 2C). Control mice (expressing DIO-GFP in dCA1) showed impaired memory linking: freezing in Context A was significantly lower than in shock-paired Context B and comparable to neutral Context C.In contrast, opto-D1 activation restored normal memory linking, as freezing in Context A was significantly higher than in Context C and comparable to Context B (Fig. 2C).These results indicate that activating D1 receptors in dCA1 during Context A exploration rescues LC–dCA1 inhibition–induced memory-linking deficits.
To test whether opto-D1 rescues memory linking by restoring dCA1 excitability, neuronal firing was examined 5 hours later.Compared with GFP controls, opto-D1 activation increased firing frequency of dCA1 neurons without altering resting membrane potential, input resistance, minimal current for excitation, after-hyperpolarization, or action potential threshold (Fig. 2D).This suggests that opto-D1 enhances excitability to rescue memory linking without affecting fundamental membrane properties or action potential generation.
Figure 2. Activation of optogenetic D1 receptors in dCA1 rescues memory-linking deficits caused by LC–dCA1 inhibition.
Photosensitive μ-Opioid Receptor (opto-MOR)
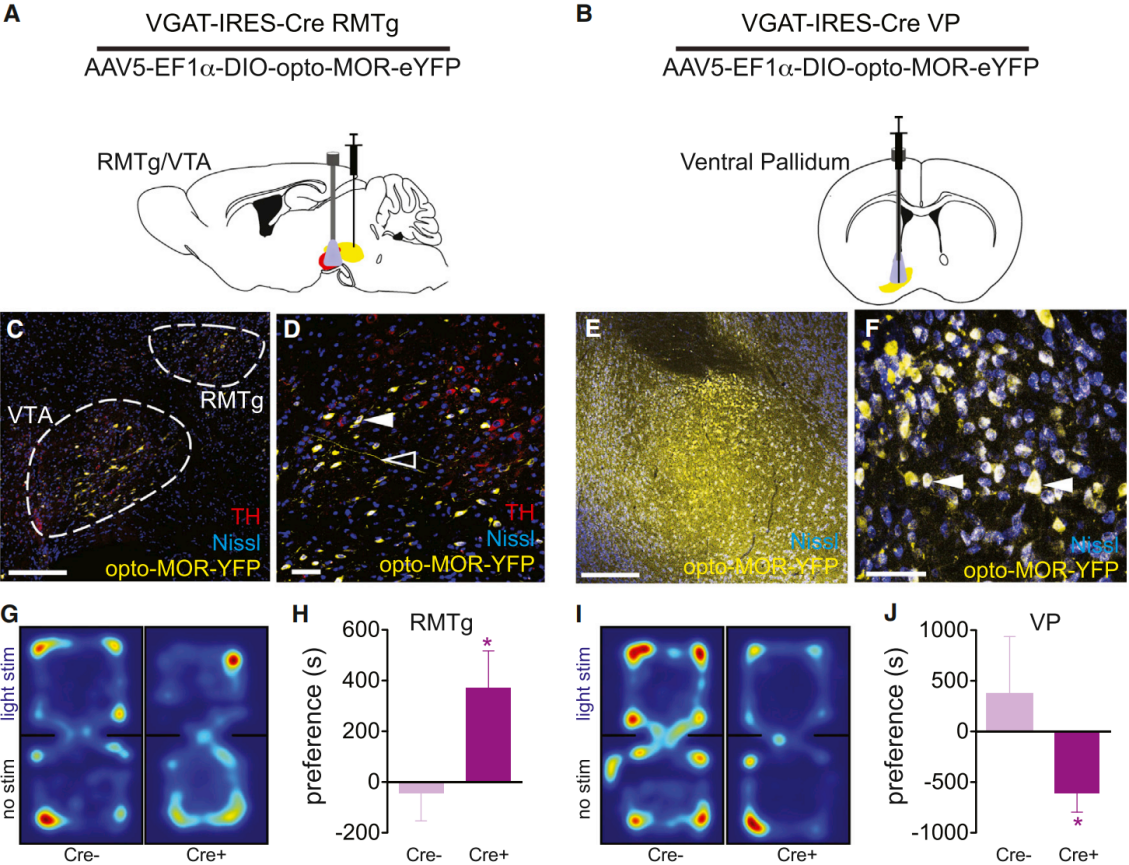
On May 20, 2015, researchers from the University of Washington published a study in Neuron titled “Spatiotemporal control of opioid signaling and behavior”.They developed and characterized a photosensitive μ-opioid receptor (opto-MOR) by replacing the intracellular loops and C-terminal domain of rat rhodopsin with the corresponding regions of the μ-opioid receptor (MOPR). In vitro, opto-MOR successfully mimicked the classical signaling pathways of MOPR, including inhibition of cAMP production, activation of MAPK (pERK), and coupling to GIRK channels, while displaying internalization and desensitization kinetics similar to native MOPR. This tool provides a novel means for spatiotemporal precision control of opioid signaling and for dissecting related neural circuits. To demonstrate the utility of opto-MOR in studying opioid-related circuits and behaviors, AAV5-EF1a-DIO-opto-MOR-YFP was injected into the rostromedial tegmental nucleus (RMTg) and ventral pallidum (VP) of vGAT-IRES-Cre mice (Fig. 3A–F). Three weeks later, optical fibers were implanted above the VTA and VP.Mice were placed in a black chamber with two compartments and allowed to freely explore. When they entered the “light stimulation” side, they received 473 nm blue light stimulation (10 mW) (Fig. 3G, 3I).Real-time place preference tests were used to evaluate behavioral responses. Results showed that opto-MOR activation in the RMTg induced a significant preference for the stimulation side compared with littermate controls (Fig. 3G–H). In contrast, opto-MOR activation in the VP produced a significant place aversion relative to controls (Fig. 3I–J).
Figure 3. Light stimulation of opto-MOR elicits MOR-like behavioral phenotypes.
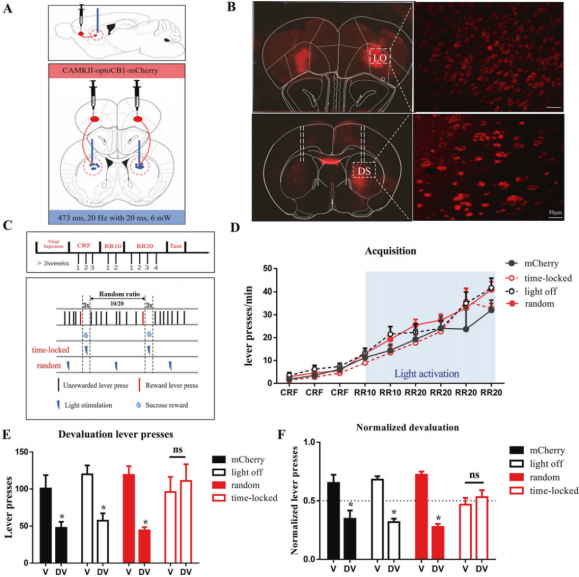
In vitro experiments showed that light activation of opto-CB1R triggered classical CB1R signaling—including inhibition of cAMP and activation of the MAPK pathway, without affecting cGMP or IP1—and rapidly (within seconds) suppressed excitatory postsynaptic currents (EPSCs) in corticostriatal glutamatergic neurons.
In vivo experiments demonstrated that brief (3-second), temporally locked activation of opto-CB1R in corticostriatal glutamatergic neurons (synchronized with reward delivery) reduced sensitivity to outcome devaluation, thereby suppressing goal-directed behavior and promoting habit formation (Fig. 4).
In contrast, prolonged (≈30-minute) activation of opto-CB1R in hippocampal astrocytes impaired working memory, whereas short activations (10 or 30 seconds) had no effect.
Notably, administration of an adenosine A2A receptor antagonist (KW6002) reversed these cognitive impairments, suggesting that the A2A receptor may be a potential therapeutic target for improving CB1R-related cognitive dysfunction.
Figure 4. Activation of opto-CB1R in corticostriatal glutamatergic neurons promotes habit formation.
Photosensitive Adenosine A2A Receptor (opto-A2AR)
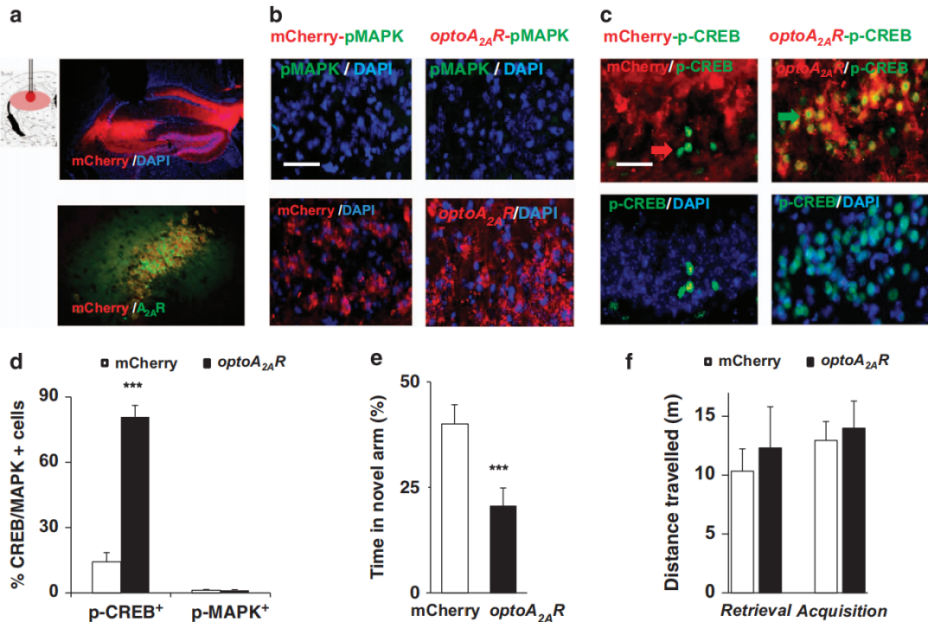
On February 17, 2015, researchers from Wenzhou Medical University published a study in Molecular Psychiatry titled“Optogenetic activation of intracellular adenosine A2A receptor signaling in the hippocampus is sufficient to trigger CREB phosphorylation and impair memory.” To test whether abnormal activation of hippocampal adenosine A2A receptors (A2ARs) is sufficient to cause memory impairment, the team developed a photosensitive adenosine A2A receptor (opto-A2AR).This receptor was engineered by replacing the intracellular loops (1/2/3) and C-terminal domain of bovine rhodopsin with the corresponding sequences of human A2AR, thereby retaining light sensitivity while mediating A2AR-specific signaling.
In vitro experiments showed that light activation of opto-A2AR selectively activated the cAMP/PKA and MAPK pathways (without affecting cGMP or IP1), producing effects consistent with the A2AR agonist CGS21680. In vivo, after injecting AAV5-CaMKIIα-opto-A2AR-mCherry into the hippocampus for two weeks (Fig. 5A), opto-A2AR activation induced a robust increase in CREB phosphorylation (p-CREB) (Fig. 5B–D) and significantly impaired spatial memory, reducing new-arm exploration in the Y-maze by 50% (Fig. 5E–F).When activated in the nucleus accumbens (NAc), opto-A2AR stimulation elevated MAPK phosphorylation (p-MAPK) and enhanced locomotor activity, increasing total movement distance by 83%.
Figure 5. Light activation of hippocampal opto-A2AR triggers CREB phosphorylation and memory impairment.
Limitations
Photosensitive receptor tools cannot fully replicate all the functions of their endogenous counterparts, which restricts comprehensive analysis of complex signaling networks.Depending on the brain region and experimental objectives, specific optimal parameters—including light stimulation duration, frequency, and intensity—are required. During long-term or high-frequency light stimulation, receptor internalization and desensitization may occur, leading to temporary loss of receptor function and significantly constraining the experimental time window. Therefore, experimental design must carefully account for these kinetic properties, allowing sufficient receptor recovery time to maintain relatively stable receptor function and to avoid drawing incorrect conclusions due to receptor inactivation.
BrainCase offers a comprehensive portfolio of vectors for opto-D1, opto-MOR, opto-CB1R, and opto-A2AR light-sensitive receptors. In addition, we specialize in tailored vector design and high-quality packaging services to meet your specific research needs. For further information or to discuss a customized solution, please feel free to contact us at bd@ebraincase.com — we would be delighted to support your work.
References
Gunaydin LA, Grosenick L, Finkelstein JC, et al. Natural neural projection dynamics underlying social behavior. Cell. 2014;157(7):1535-1551.
Chowdhury A, Luchetti A, Fernandes G, et al. A locus coeruleus-dorsal CA1 dopaminergic circuit modulates memory linking. Neuron. 2022;110(20):3374-3388.e8.
Siuda ER, Copits BA, Schmidt MJ, et al. Spatiotemporal control of opioid signaling and behavior. Neuron. 2015;86(4):923-935.
Shang H, Li P, Lin X, et al. Neuronal and astrocytic CB1R signaling differentially modulates goal-directed behavior and working memory by distinct temporal mechanisms. Neuropsychopharmacology. 2023;48(10):1520-1531.
Li P, Rial D, Canas PM, et al. Optogenetic activation of intracellular adenosine A2A receptor signaling in the hippocampus is sufficient to trigger CREB phosphorylation and impair memory. Mol Psychiatry. 2015;20(11):1339-1349.
Service Type :
Select the service you'd like to purchase.
Order Information(Premade-AAVs)
Please provide us some information about the service you'd like to order.
Order Information(Custom AAV/Lentivirus)
Please provide us some information about the service you'd like to order.
Order Information(Others)
Please provide us some information about the service you'd like to order.