Cellular Autophagy: Main types, Mechanisms, Biomarkers, and Fluorescent Reporter Tools
Release time:2025-09-09 13:59:24
The meaning of cellular autophagy:
Autophagy is a highly conserved physiological process of degradation and recycling within eukaryotic cells. It was first proposed in 1963 by Belgian scientist Christian de Duve. Its core function is to degrade damaged organelles, misfolded proteins, or invading pathogens within the cell, and to recycle the resulting products (such as amino acids, fatty acids, and nucleotides). This process helps maintain intracellular homeostasis and enables cells to cope with external stresses (e.g., nutrient deprivation, oxidative stress). Autophagy is vividly referred to as the cell’s “waste disposal plant” and “emergency granary,” playing a crucial role in cell survival, development, and disease progression.
Main types of cellular autophagy
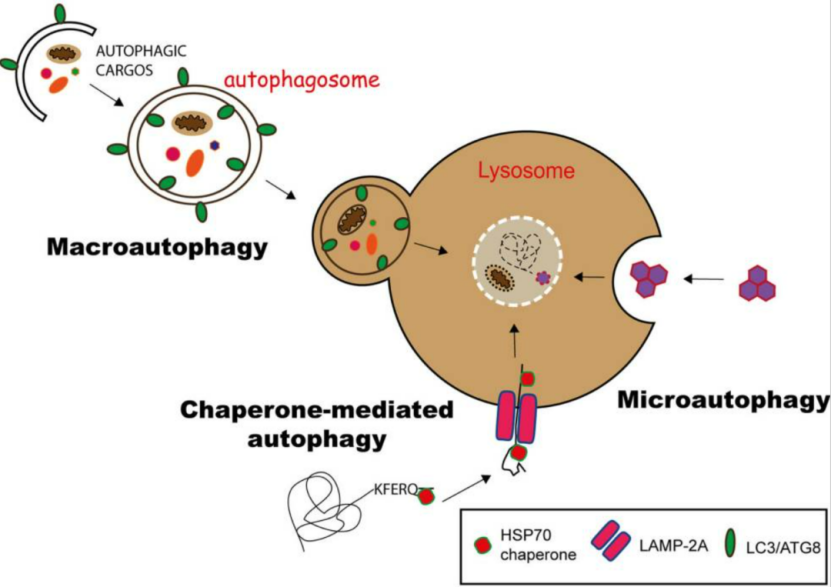
Based on differences in the nature of degraded substrates, the mode of sequestration, and the underlying mechanisms, cellular autophagy can be divided into three types: 1.Macroautophagy: The most extensively studied and predominant type. It involves the formation of a double-membrane structure called the autophagosome, which engulfs damaged organelles (such as abnormal mitochondria) or large aggregates of misfolded proteins. The autophagosome then fuses with the lysosome to form the autolysosome, where both the autophagosome membrane and its contents are completely degraded.
2. Microautophagy: This process does not require an autophagosome. Instead, the lysosomal membrane directly invaginates and folds inward to engulf small proteins or fragments of organelles, which are subsequently degraded by lysosomal enzymes. Due to the limited capacity of lysosomal membrane invagination, microautophagy primarily handles small molecules or fragments.
3. Chaperone-mediated autophagy (CMA): A highly selective form of autophagy. Specific chaperone proteins (such as Hsc70) recognize proteins containing particular amino acid motifs (KFERQ-like sequences) and directly “escort” them to the lysosomal membrane for translocation and degradation. This process does not involve the formation of autophagosomes.
Figure 1. Schematic illustration of the three autophagy pathways
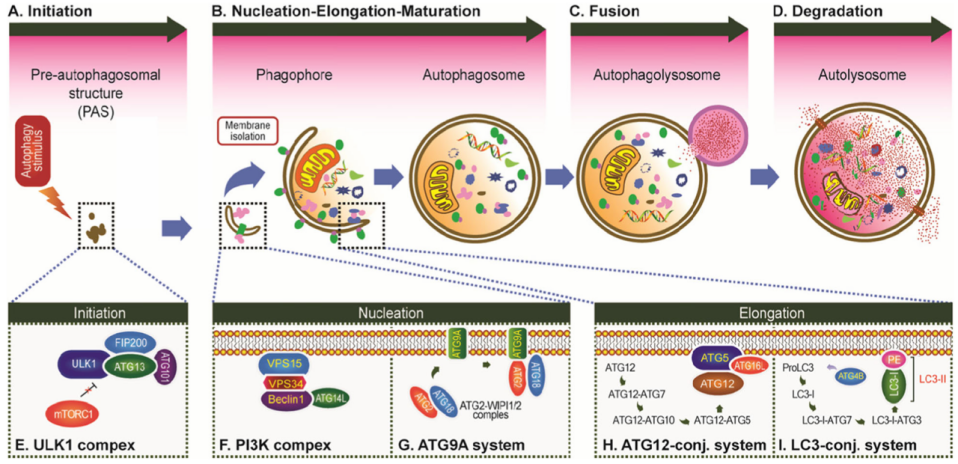
Cellular Autophagy Process (using the most common type, macroautophagy, as an example)
The core of autophagy is membrane reorganization within the cell. Through this process, a double-membrane vesicleforms to enclose random or specific substrates, which then fuses with a lysosome to generate an autolysosome, ultimately leading to substrate degradation. The process of autophagy can be broadly divided into four stages: 1.Initiation of autophagy When cells sense stress signals (e.g., nutrient deprivation, hypoxia, or organelle damage), autophagy-related genes (ATG family) are activated. Among these, the ULK complex (a key autophagy-initiating complex) and the Beclin-1 complex (which regulates membrane nucleation) become activated.
2.Formation of the isolation membrane and autophagosome Under the regulation of the Beclin-1 complex and other factors, membrane structures within the cell gradually aggregate and elongate to form a double-membrane precursor, known as the isolation membrane (phagophore). This structure expands and sequesters materials targeted for degradation (e.g., damaged mitochondria, protein aggregates), ultimately forming a mature autophagosome.
3.Fusion of the autophagosome with the lysosome The mature autophagosome is transported toward the lysosome, an organelle rich in hydrolytic enzymes. The two fuse to form an autolysosome, exposing the autophagosome contents to the lysosomal enzymatic environment.
4.Degradation within the autolysosome Hydrolytic enzymes (such as proteases and nucleases) within the autolysosome thoroughly break down the sequestered materials into small molecules including amino acids, fatty acids, and nucleotides, which are then recycled for cellular use.
Figure 2. The process of autophagy
Autophagy Marker LC3
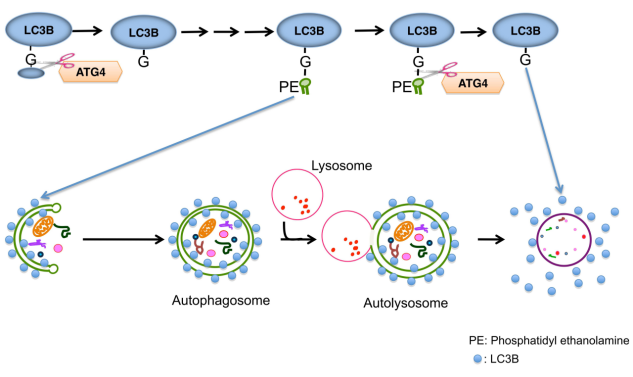
LC3 protein, also known as microtubule-associated protein 1 light chain 3, is one of the most important markers in the autophagy process. In mammals, LC3 exists in three isoforms: LC3A, LC3B, and LC3C, among which LC3B is the most widely used. Under normal conditions, LC3 exists in the cytoplasm in its soluble form, LC3-I. Upon autophagy activation, LC3-I undergoes modification through a ubiquitin-like conjugation system and is eventually conjugated to phosphatidylethanolamine (PE), converting into the membrane-bound form LC3-II. LC3-II anchors to both the inner and outer membranes of autophagosomes, assisting in autophagosome membrane expansion and sequestration of damaged organelles or proteins, while also mediating autophagosome–lysosome fusion. The level of LC3-II (e.g., detected by Western blot) or the number of intracellular LC3 puncta is commonly used to evaluate autophagic activity.
Figure 3. The role of LC3 in autophagy.
Autophagy Single-Fluorescent Reporter Virus
Single-fluorescent reporter viruses for autophagy, such as GFP-LC3, can be used to monitor autophagic activity. When autophagy is absent, GFP-LC3 is diffusely distributed throughout the cytoplasm. Once autophagy is induced, the GFP-LC3 fusion protein aggregates and translocates to autophagosome membranes. Under a fluorescence microscope, the number of fluorescent puncta corresponds to the number of autophagosomes, thereby reflecting the level of autophagic activity.
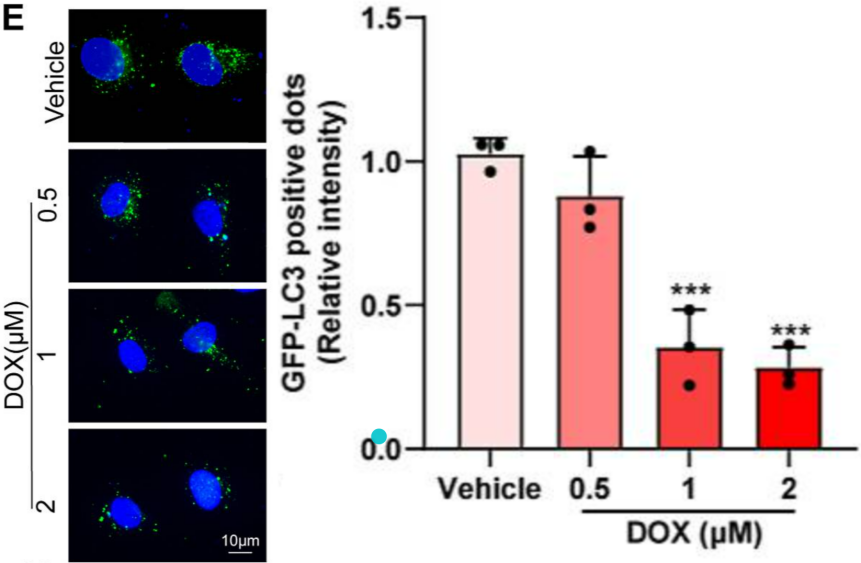
Example Demonstration 1
CMECs were transduced with GFP-LC3 via lentivirus for 48 hours, followed by treatment with different concentrations of DOX. Fluorescent puncta were then observed and quantified under a fluorescence microscope. The results showed that with increasing DOX concentrations, cells exhibited dose-dependent autophagy inhibition, indicating that DOX suppresses autophagy.
Figure 4. DOX induces dose-dependent autophagy impairment in cells.
Autophagy Dual-Fluorescent Reporter Virus The mRFP-GFP-LC3 virus can be used to label and track LC3 as well as changes in autophagic flux. GFP is sensitive to acidic environments; in the acidic lumen of autolysosomes, the decrease in pH quenches GFP fluorescence. In contrast, mRFP is a stable fluorescent protein unaffected by such conditions. By observing the color changes of intracellular fluorescent puncta: newly formed autophagosomes appear as yellow puncta (GFP+/mRFP+), while autolysosomes after fusion with lysosomes appear as red puncta only (GFP-/mRFP+). This allows for a direct and dynamic assessment of autophagic flux.
Example Demonstration 2
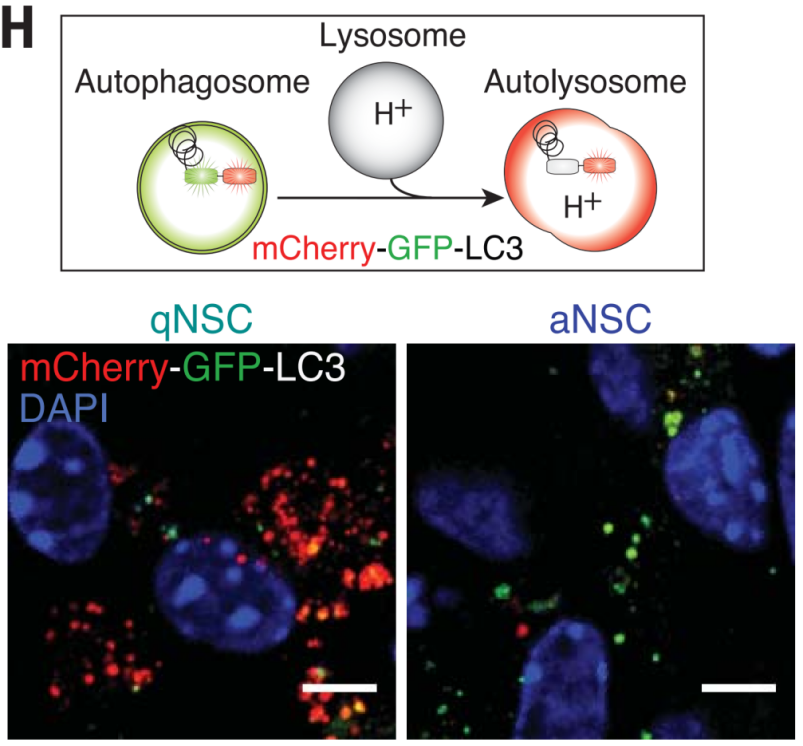
1.NSCs are a collective term for stem cells in the neurogenic microenvironment of the brain that can self-renew and differentiate into neural cells. They consist of quiescent NSCs (qNSCs) and activated NSCs (aNSCs), which play different roles. Transcriptomic analysis has shown differences in the protein homeostasis networks of qNSCs and aNSCs: aNSCs contain active proteasomes, while qNSCs contain large lysosomes. In primary cultured qNSCs and aNSCs from 3-month-old mice expressing mCherry-GFP-LC3, autophagosomes display both GFP and mCherry fluorescence (yellow-green), whereas autolysosomes display only mCherry fluorescence (red), as GFP is denatured and quenched in the acidic lysosomal environment. The results showed that qNSCs contain many large lysosomes that have already fused with autophagosomes to form autolysosomes, but their contents have not yet been degraded.
Figure 5. Autophagy in qNSCs and aNSCs.
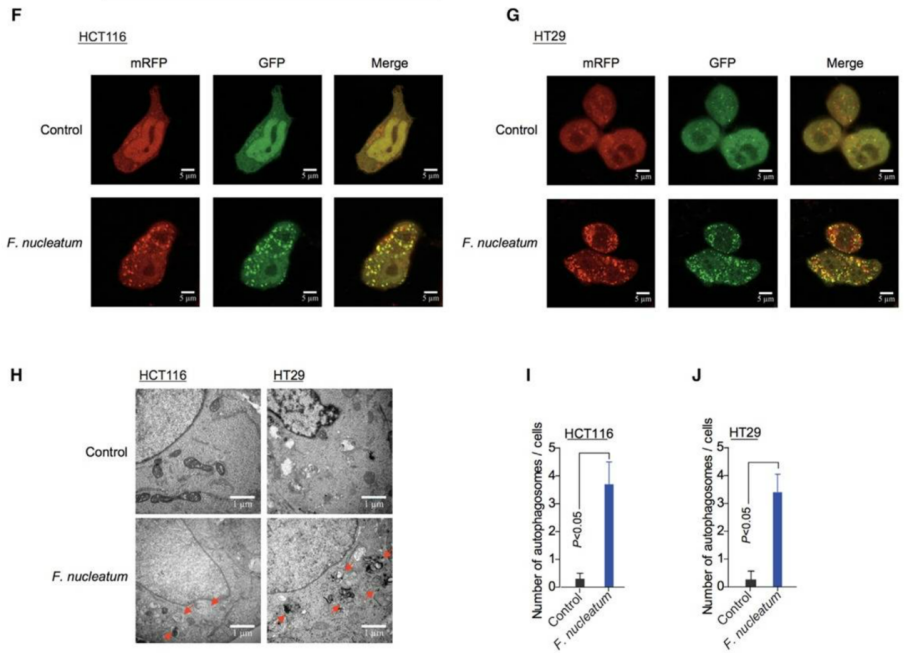
HCT116 and HT29 cells stably expressing the mRFP-EGFP-LC3 construct were co-cultured with Fusobacterium nucleatum (F. nucleatum). The results showed that after F. nucleatum induction, the number of red and green fluorescent puncta increased in both HCT116 and HT29 cells. Transmission electron microscopy further revealed enhanced formation of autophagic vesicles in HCT116 and HT29 cells co-cultured with F. nucleatum. These findings indicate that F. nucleatum can activate autophagy pathways in cells.
Figure 6. Induction of autophagy by F. nucleatum in cells.
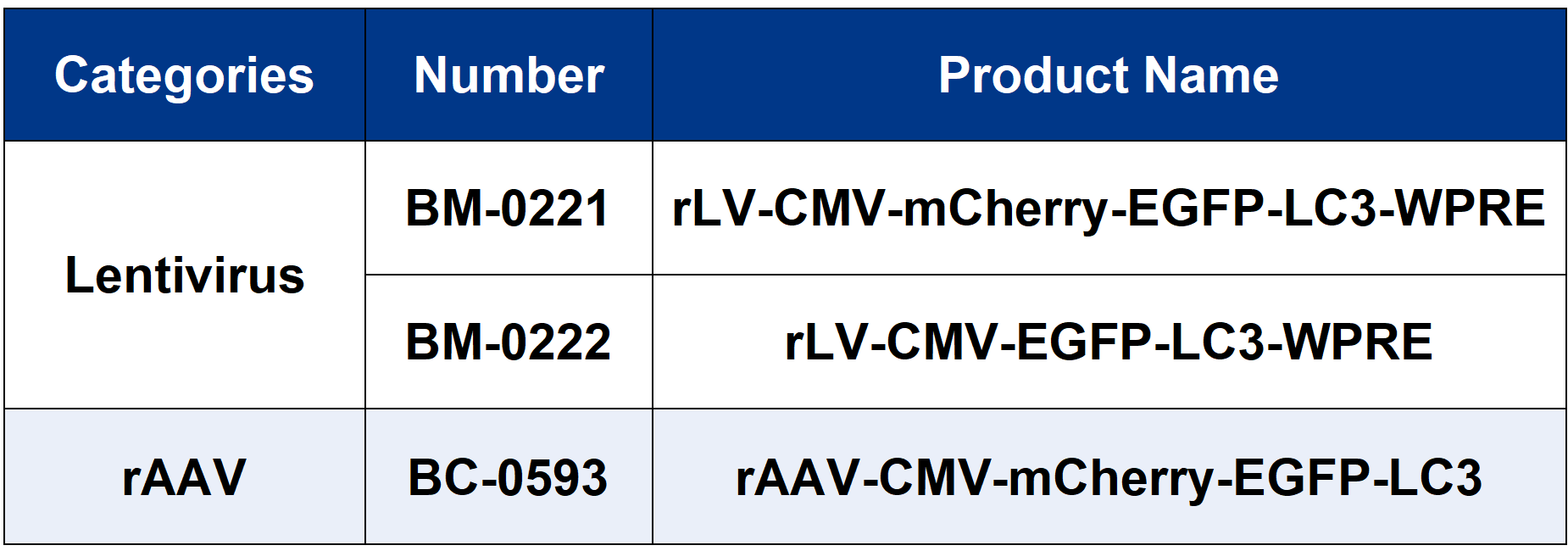
Brain Case provides related autophagy tool viruses. For more information,
please don’t hesitate to reach out to us at bd@ebraincase.com
Related References:
1. Durães FV, Niven J, Dubrot J, Hugues S, Gannagé M. Macroautophagy in endogenous processing of self- and pathogen-derived antigens for MHC class II presentation. Front Immunol. 2015 Sep 22;6:459. 2. Li X, He S, Ma B. Autophagy and autophagy-related proteins in cancer. Mol Cancer. 2020 Jan 22;19(1):12. 3. Pan JA, Zhang H, Lin H, Gao L, Zhang HL, Zhang JF, Wang CQ, Gu J. Irisin ameliorates doxorubicin-induced cardiac perivascular fibrosis through inhibiting endothelial-to-mesenchymal transition by regulating ROS accumulation and autophagy disorder in endothelial cells. Redox Biol. 2021 Oct;46:102120. 4. Leeman DS, Hebestreit K, Ruetz T, Webb AE, McKay A, Pollina EA, Dulken BW, Zhao X, Yeo RW, Ho TT, Mahmoudi S, Devarajan K, Passegué E, Rando TA, Frydman J, Brunet A. Lysosome activation clears aggregates and enhances quiescent neural stem cell activation during aging. Science. 2018 Mar 16;359(6381):1277-1283. 5. Yu T, Guo F, Yu Y, Sun T, Ma D, Han J, Qian Y, Kryczek I, Sun D, Nagarsheth N, Chen Y, Chen H, Hong J, Zou W, Fang JY. Fusobacterium nucleatum promotes chemoresistance to colorectal cancer by modulating autophagy. Cell. 2017 Jul 27;170(3):548-563.e16.
Service Type :
Select the service you'd like to purchase.
Order Information(Premade-AAVs)
Please provide us some information about the service you'd like to order.
Order Information(Custom AAV/Lentivirus)
Please provide us some information about the service you'd like to order.
Order Information(Others)
Please provide us some information about the service you'd like to order.