Science: Yulong Li’s Team Develops HaloDA1.0 Far-Red Dopamine Sensor, Enabling Simultaneous Monitoring of Multiple Neurochemical Signals
Release time:2025-06-20 15:46:09
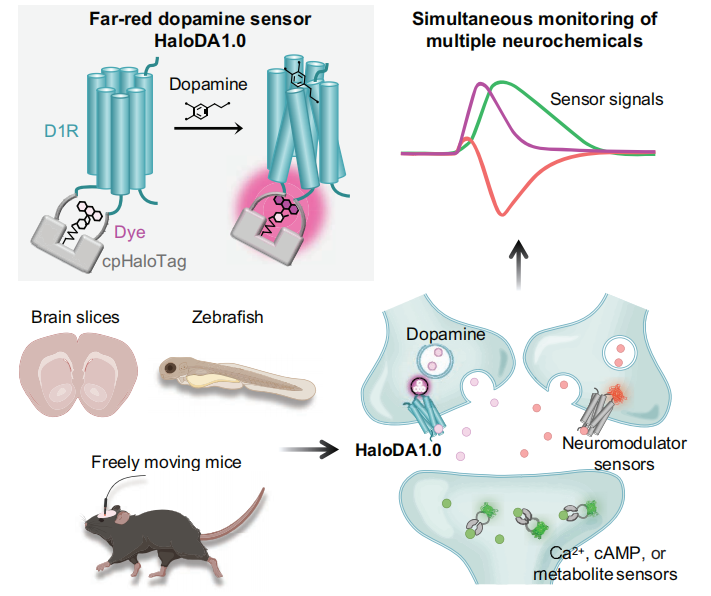
Neurochemical signals—including neurotransmitters, neuromodulators, and intracellular signaling molecules—play essential roles in regulating various brain functions and mediating neurological diseases. Dopamine (DA), one of the most critical neuromodulators, interacts in complex ways with other neuromodulators such as acetylcholine (ACh) and endocannabinoids (eCBs), as well as with intracellular messengers like cyclic AMP (cAMP) and calcium ions (Ca²⁺). Deciphering these intricate neurochemical networks is vital for understanding the neural mechanisms underlying behavior and related disorders. However, current genetically encoded sensors are limited to green and red spectral ranges, constraining the real-time simultaneous detection of multiple neurochemical signals. Thus, there is an urgent need to expand the spectral coverage of neuromodulator sensors into the far-red and near-infrared (NIR) wavelengths (>650 nm).
On June 5, 2025, Yulong Li’s lab at Peking University published a study titled “In vivo multiplex imaging of dynamic neurochemical networks with designed far-red dopamine sensors” in Science. The team developed a novel far-red dopamine sensor, HaloDA1.0, which combines a cpHaloTag–chemical dye system with a G-protein-coupled receptor activation-based (GRAB) strategy. HaloDA1.0 exhibits high sensitivity to dopamine and enables multiplexed neurochemical imaging across diverse biological systems. Using this sensor, the researchers successfully achieved dynamic, simultaneous monitoring of DA, ACh, and eCBs, revealing their complex interactions under various physiological and behavioral conditions.
Development and In Vitro Characterization of the Far-Red DA Sensor
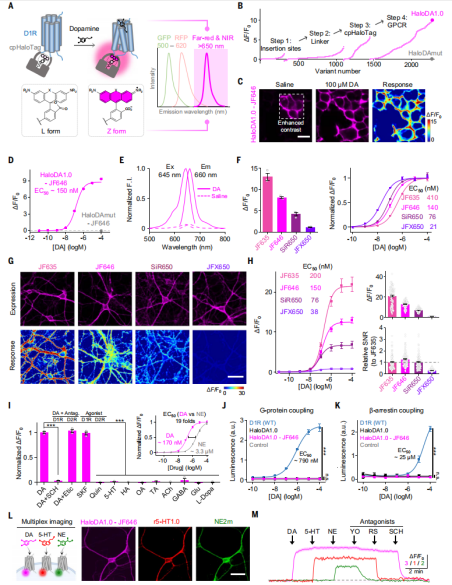
The researchers used the human D1 receptor (D1R) as the DA sensing module, due to its superior membrane trafficking properties compared to other DA receptors. They replaced the third intracellular loop (ICL3) of D1R with an optimized circularly permuted HaloTag protein (cpHaloTag), and labeled the chimeric variants with far-red fluorescent dyes conjugated to HaloTag ligands, which can form covalent bonds with cpHaloTag. Upon binding of DA to D1R, a conformational change is induced in the receptor, which subsequently triggers a conformational shift in cpHaloTag, altering the conjugated dye from a non-fluorescent (L) state to a fluorescent (Z) state, ultimately enhancing fluorescence (Fig. 1A).
The researchers primarily used JF646 as the far-red dye and systematically optimized the insertion site of cpHaloTag, the linker sequences, and critical residues in both cpHaloTag and D1R. After screening over 2,000 variants, they identified the one with the strongest response and named it HaloDA1.0. Additionally, they generated a DA-insensitive sensor (HaloDAmut) by mutating key residues in the ligand-binding pocket, which served as a negative control (Fig. 1B).
They confirmed that the JF646-conjugated HaloDA1.0 sensor (HaloDA1.0-JF646) localized to the plasma membrane when expressed in HEK293T cells (Fig. 1C), and produced a robust, rapid fluorescence increase upon ligand application. The half-maximal effective concentration (EC50) was 150 nM, with a maximum ΔF/F₀ of approximately 900% (Fig. 1D). Using one-photon excitation, they confirmed that HaloDA1.0-JF646 operates in the far-red range, with an excitation peak at 645 nm and an emission peak at 660 nm (Fig. 1E). Since the structure and properties of chemical dyes vary, these factors influenced HaloDA1.0’s performance. When labeled with different far-red dyes, the peak ΔF/F₀ response ranged from 110% to 1300%, and EC50 values ranged from 27 nM to 410 nM (Fig. 1F), with significant differences in fluorescence brightness and labeling efficiency. Regardless of the dye used, the DA-insensitive HaloDAmut sensor showed no fluorescence increase (Fig. 1D). When applied to cultured neurons, HaloDA1.0 labeled with dyes showed results consistent with HEK293T cells, with all four tested dyes showing a similar rank order of peak response and DA affinity (Fig. 1G and 1H). These findings demonstrate that the characteristics of HaloDA1.0—including its spectrum, brightness, ligand responsiveness, and ligand affinity—can be finely tuned by selecting specific chemical dyes.
Moreover, HaloDA1.0 exhibited high sensitivity and specificity for DA. The DA-induced fluorescence enhancement could be blocked by co-application of SCH, a D1R-specific antagonist, but not by Etic, a D2R-specific antagonist (Fig. 1I). To examine whether HaloDA1.0 couples to downstream intracellular signaling pathways, the researchers used luciferase complementation and Tango assays to assess Gs and β-arrestin signaling. HaloDA1.0 only weakly activated these pathways, in contrast to wild-type D1R (WT), which robustly activated both in a dose-dependent manner (Fig. 1J and 1K).
Finally, in cultured neurons, the researchers co-expressed the far-red HaloDA1.0-JF646 sensor, the red fluorescent serotonin sensor (r5-HT1.0), and the green fluorescent norepinephrine sensor (NE2m). They found that all three sensors could be expressed in the same neuron and that their respective fluorescence signals could be selectively activated and blocked by applying their corresponding agonists and antagonists. This enabled real-time, simultaneous monitoring of dynamic signaling from multiple neuromodulators (Fig. 1L and 1M).
Figure 1. Construction and characterization of the HaloDA1.0 dopamine sensor
HaloDA1.0 Sensor Enables Multiplex Imaging in Acute Brain Slices
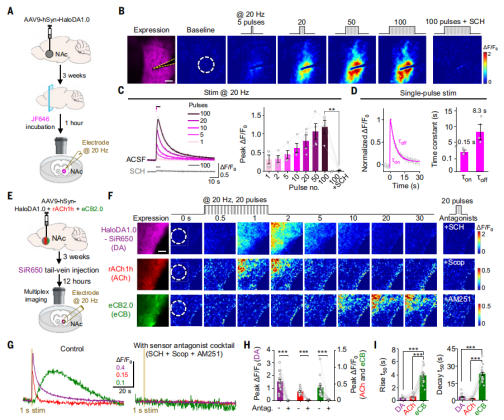
To assess whether HaloDA1.0 can detect endogenous DA release, researchers injected adeno-associated virus (AAV) encoding HaloDA1.0 into the nucleus accumbens (NAc) of mice. Three weeks after expression, acute brain slices were prepared and incubated with JF646 for one hour to label the sensor (Fig. 2A). They found that local electrical stimulation at 20 Hz induced a strong and transient fluorescence increase. The response magnitude correlated with the number of stimulation pulses, and the fluorescence signal induced by endogenous DA release could be blocked by application of the D1R antagonist SCH (Fig. 2B and 2C). Furthermore, HaloDA1.0 exhibited high sensitivity, capable of detecting DA release triggered by a single electrical pulse. The fluorescence signal showed an average rise time constant of 150 ms and a decay time constant of 8.3 s (Fig. 2D).
In addition to DA, the NAc is known to release various other neuromodulators such as ACh and eCBs. The researchers injected a viral mixture into the NAc that expressed HaloDA1.0 (subsequently labeled with the far-red dye SiR650 via tail vein injection), rACh1h (a red fluorescent ACh sensor), and eCB2.0(Fig. 2E). All three sensors responded to 20 Hz field stimulation with significant fluorescence increases. The signal from each sensor could be specifically blocked by applying its corresponding antagonist, confirming specificity (Fig. 2F–2H).
Compared to the DA and ACh signals, the eCB signal showed slower rise and decay kinetics, with eCB release occurring after the stimulation ended (Fig. 2I). This finding is consistent with previous electrophysiological studies, which have shown that eCBs require time to be synthesized before release, whereas DA and ACh are released directly from vesicles upon stimulation. These results demonstrate that HaloDA1.0 is well-suited for multiplex neurochemical imaging.
Figure 2. Multiplex imaging using HaloDA1.0 in acute brain slices
HaloDA1.0 Sensor Enables Multiplex Imaging in Zebrafish Larvae
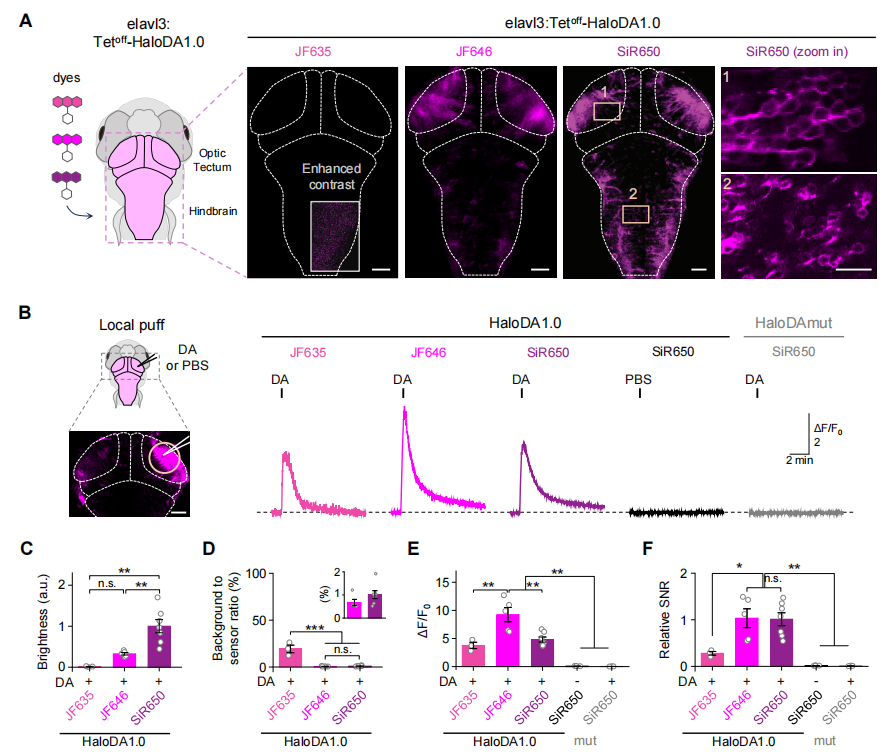
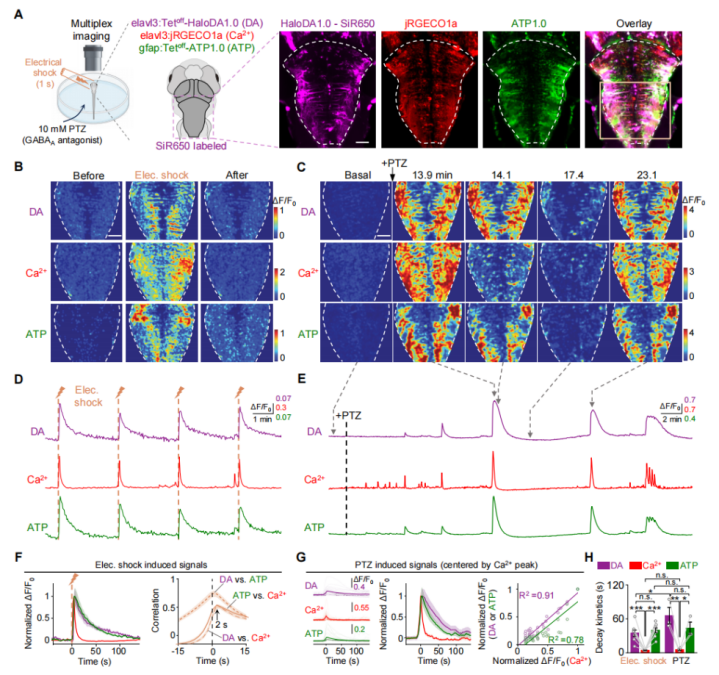
To determine whether HaloDA1.0 can be used to monitor DA dynamics in vivo, the sensor was transiently expressed in neurons of zebrafish larvae. The sensor was subsequently labeled with three different far-red dyes—JF635, JF646, and SiR650. Among them, SiR650-labeled HaloDA1.0 exhibited the strongest baseline fluorescence (Fig. 3A–3C). Both JF646 and SiR650 showed relatively low background staining, approximately 1% of the sensor’s fluorescence signal (Fig. 3D). In addition, local field stimulation induced a rapid and robust transient fluorescence increase, with the JF646-labeled sensor showing the largest response amplitude (Fig. 3E). Sensors labeled with SiR650 and JF646 displayed comparable signal-to-noise ratios (SNR), both superior to the JF635-labeled variant (Fig. 3F).
Next, three-color in vivo imaging was performed in zebrafish larvae. HaloDA1.0 was transiently expressed in neurons, along with the red fluorescent calcium sensor (jRGECO1a) and the green fluorescent ATP sensor (ATP1.0). In motor neurons, HaloDA1.0 was labeled with SiR650. Upon mild electrical stimulation, fluorescence increases were observed from all three sensors. The DA and ATP signals exhibited similar kinetics, both decaying more slowly than the neuronal calcium signal (Fig. 4). These results demonstrate that HaloDA1.0 reliably detects DA release in vivo and is well-suited for three-color imaging in the zebrafish brain.
Figure 3. Performance of the HaloDA1.0 sensor in zebrafish
Figure 4. Multiplex imaging using HaloDA1.0 in zebrafish
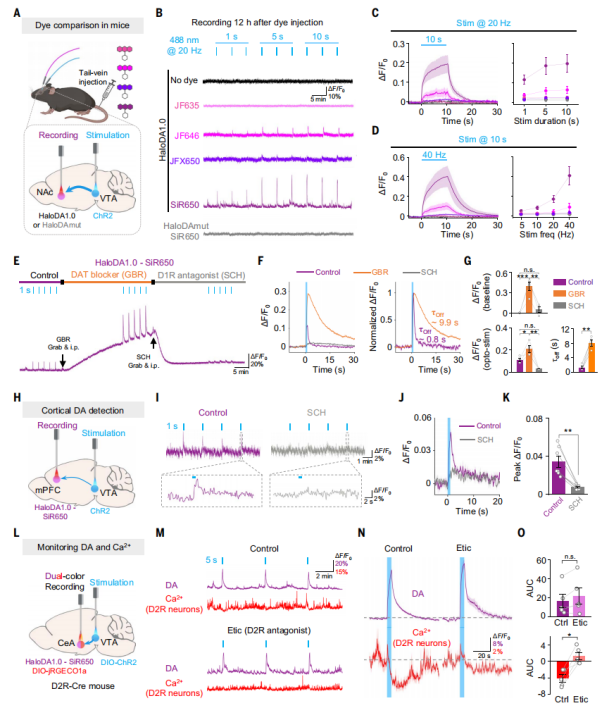
HaloDA1.0 Detects Optogenetically Evoked Dopamine Release in Freely Moving Mice
Channelrhodopsin-2 (ChR2) viral vectors for optogenetics were expressed in the ventral tegmental area (VTA) of mice, while the HaloDA1.0 sensor was expressed in the nucleus NAc, which receives dense dopaminergic projections from the VTA (Fig. 5A). Different dyes were injected intravenously to label the sensor, and fiber photometry recordings were performed 12 hours later. The results showed that optogenetic stimulation of the VTA induced a fluorescence increase of HaloDA1.0 labeled with JF646, whereas sensors labeled with JF635 or JFX650 showed no detectable changes (Fig. 5B–5D). Consistent with results obtained in zebrafish, the HaloDA1.0 sensor labeled with SiR650 exhibited a much stronger response. Moreover, after intraperitoneal injection of the dopamine transporter (DAT) blocker GBR, the baseline fluorescence of SiR650-labeled HaloDA1.0 slowly increased, and both the amplitude and decay time of the optically evoked responses were enhanced (Fig. 5E–5G). Treatment with the D1R antagonist SCH eliminated both the baseline fluorescence increase and the optogenetically evoked response.
Next, the researchers tested whether HaloDA1.0 could monitor in vivo dopamine release in brain regions with sparse neuronal distribution, such as the medial prefrontal cortex (mPFC). Activation of neurons in the VTA induced transient increases of SiR650-labeled HaloDA1.0 signals in the mPFC, and these responses were blocked by SCH (Fig. 5H–5K). To test the suitability of HaloDA1.0 for dual-color recordings during optogenetic stimulation, DIO-ChR2 was expressed in dopamine neurons of D2R-Cre mice to selectively activate VTA dopaminergic neurons. HaloDA1.0 and DIO-jRGECO1a were co-expressed in the central amygdala (CeA) (Fig. 5L). Optogenetic stimulation triggered an increase in dopamine release detected by HaloDA1.0 and a decrease in Ca²⁺ signals detected by jRGECO1a in D2R-positive neurons. Treatment with the D2 receptor (D2R) antagonist Etic blocked the Ca²⁺ changes without affecting dopamine release (Fig. 5M–5O), suggesting that dopamine may inhibit the activity of D2R-positive neurons in the CeA by activating inhibitory D2R signaling pathways.
Figure 5. HaloDA1.0 detection of endogenous dopamine release in freely moving mice
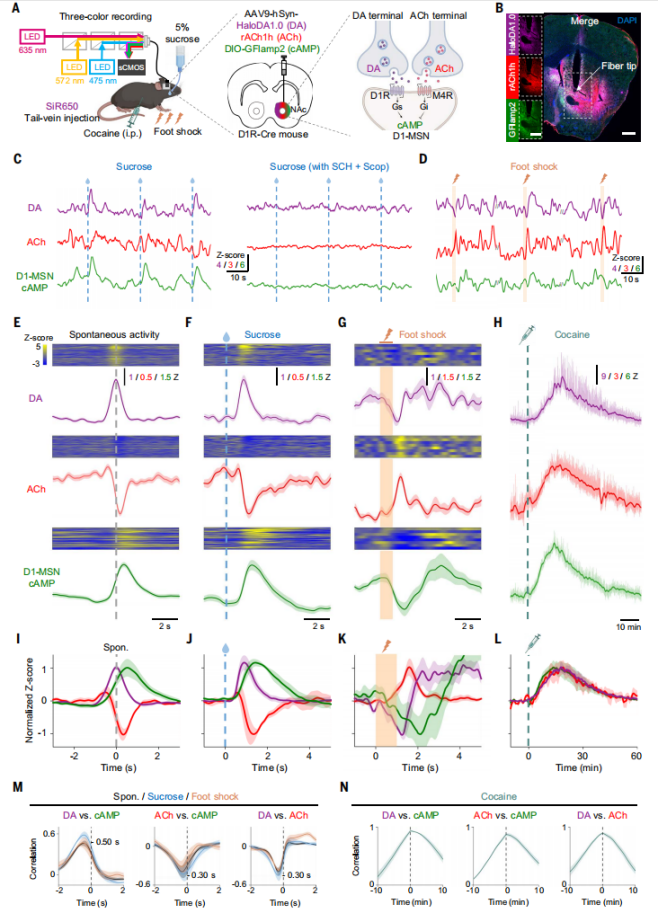
Simultaneous Monitoring of DA, ACh, and cAMP Dynamics in the Mouse Nucleus Accumbens
Viruses expressing HaloDA1.0, rACh1h, and a green fluorescent cAMP sensor (DIO-GFlamp2) were co-expressed in the NAc of D1R-Cre mice. The DA sensor was labeled with SiR650, and triple-color fiber photometry was used to simultaneously monitor in vivo dynamics of DA, ACh, and cAMP (Fig. 6A and 6B). The results showed spontaneous fluctuations in all three signals under unstimulated conditions (Fig. 6C). Centered on the peaks of spontaneous DA fluctuations, a corresponding increase in cAMP signal was observed, while the ACh signal showed a phased decrease (Fig. 6E and 6I). The peak of DA preceded the trough of the ACh signal, followed by the peak of cAMP, consistent with the requirement that DA binds to D1R to produce cAMP. Additionally, during unconditioned sucrose reward delivery, a pattern similar to spontaneous signals was observed (Fig. 6C, 6F, 6J). In contrast, during a 1-second foot shock, a distinct pattern appeared with significant changes in all three signals (Fig. 6G, 6K). Finally, correlation analysis revealed a direct positive correlation between DA and cAMP (lag of 500 ms), while negative correlations were found between ACh and cAMP and between DA and ACh (both with a lag of 300 ms) (Fig. 6M).
Figure 6. Simultaneous in vivo monitoring of DA, ACh, and cAMP dynamics
In summary, the researchers developed a chemogenetic far-red dopamine sensor, HaloDA1.0, capable of sensitive detection of dopamine both in vitro and in vivo. The HaloDA1.0 sensor demonstrated remarkable advantages across multiple biological systems, including cultured neurons, acute mouse brain slices, zebrafish, and live mice. This approach deepens our understanding of the complex interactions among various neurochemical signals and lays a foundation for further exploration of neural circuit functions as well as better comprehension of neurological and psychiatric disorders.
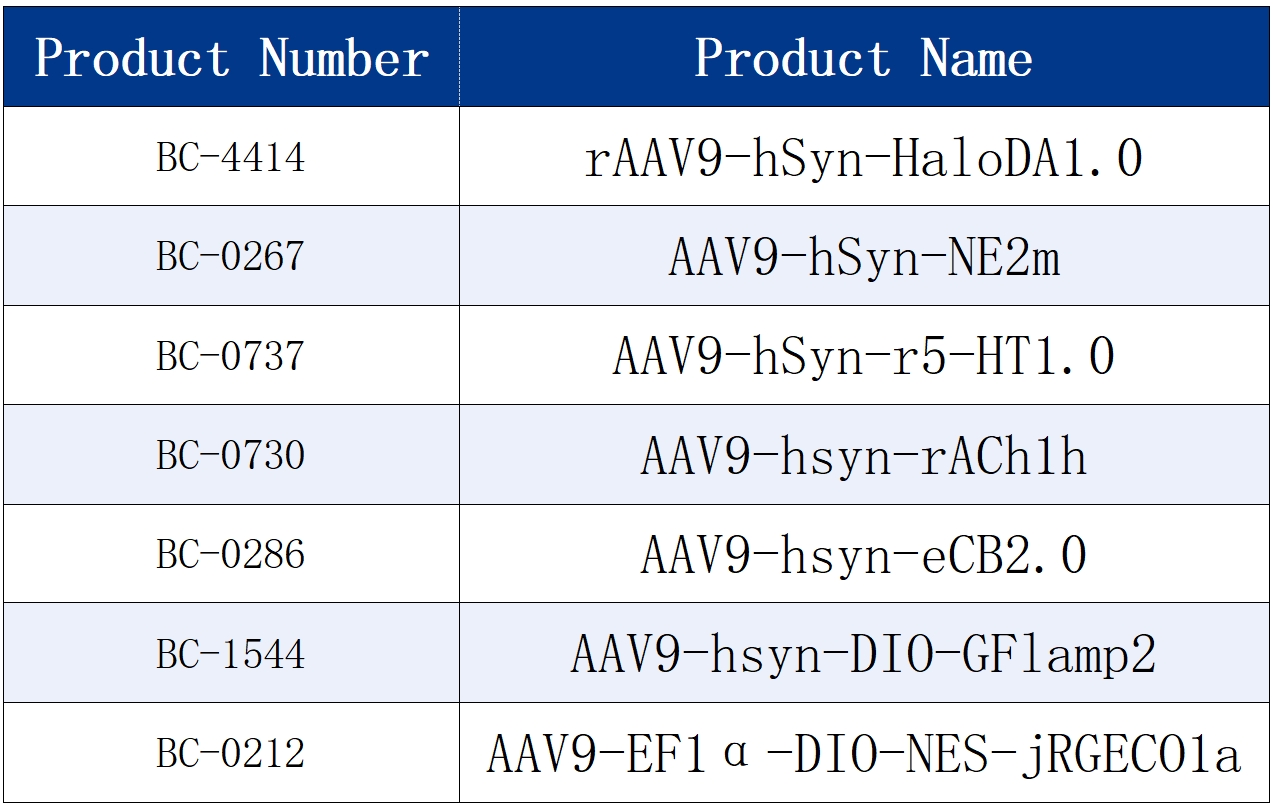
The sensor tools used in the text are all available from Brain Case.
Contact Us
Brain Case Biotechoffers various custom and viral packaging services. Please contact bd@ebraincase.com
Service Type :
Select the service you'd like to purchase.
Order Information(Premade-AAVs)
Please provide us some information about the service you'd like to order.
Order Information(Custom AAV/Lentivirus)
Please provide us some information about the service you'd like to order.
Order Information(Others)
Please provide us some information about the service you'd like to order.