Jing Wang, Zhaofa Wu, and Yulong Li’s research teams develop a novel probe to analyze the spatiotemporal dynamics and regulatory mechanisms of intracellular adenosine in vivo.
Release time:2025-06-03 15:20:13
Adenosine (Ado) is a central regulatory molecule in vital biological processes, playing multiple roles in neural plasticity, motor function, learning and memory, and sleep regulation. Its mechanisms of action differ based on its intra- or extracellular localization: extracellular adenosine (eAdo) acts like a "signaling dispatcher," regulating neurotransmission, immune response, and cardiovascular dilation via GPCRs such as A1R, A2AR, A2BR, and A3R. Its dynamic balance influences the progression of inflammation and the tumor microenvironment. In contrast, intracellular adenosine (iAdo) functions as an "invisible orchestrator" of energy metabolism. As a key product of ATP metabolism, it is involved in nucleotide synthesis, energy homeostasis, and signal transduction, with its concentration fluctuations indirectly reflecting the cellular energy state. eAdo and iAdo engage in cross-membrane communication through ENT (equilibrative nucleoside transporter) proteins, forming a regulatory network whose imbalance is closely linked to major diseases such as neurodegenerative disorders, metabolic syndrome, and cancer. However, due to limitations of traditional techniques in capturing millisecond-scale dynamics and subcellular localization of iAdo, the cellular origins, release mechanisms, and transmembrane transport processes of adenosine remain subjects of ongoing debate.
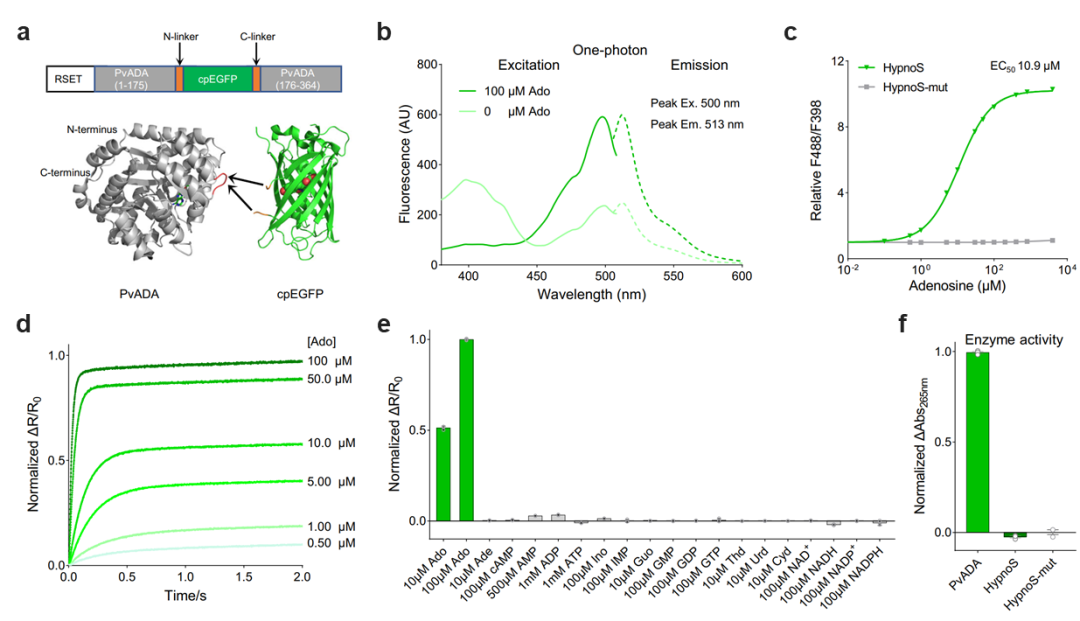
On May 7, 2025, the research team led by Jing Wang from the School of Pharmaceutical Sciences at Peking University, in collaboration with Zhaofa Wu's team from the Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, and Yulong Li's team from the School of Life Sciences at Peking University, published a study in Nature Communications titled “A high-performance fluorescent sensor spatiotemporally reveals cell-type specific regulation of intracellular adenosine in vivo.” This study successfully developed the first genetically encoded fluorescent probe for in vivo detection of intracellular adenosine, named HypnoS (Hypersensitive intracellular adenosine Sensor). Using in vivo imaging and optogenetics, the researchers explored the cell-type-specific spatiotemporal dynamics of intracellular adenosine and the underlying molecular regulatory mechanisms in living animals. The research team engineered the novel fluorescent probe HypnoS (named after “Hypnos,” the Greek god of sleep) by fusing circularly permuted enhanced green fluorescent protein (cpEGFP) with Plasmodium vivax adenosine deaminase (PvADA). After screening over 3,000 mutants, they successfully developed HypnoS, a genetically encoded sensor with outstanding features: ultra-high sensitivity (response amplitude >900%), suitable affinity (EC50 ~11 μM), sub-second kinetics, and excellent specificity (Figure 1). Notably, expression of HypnoS does not significantly interfere with normal neuronal physiology, making it an ideal tool for monitoring intracellular adenosine dynamics under both physiological and pathological conditions.
Figure 1. Development and in vitro characterization of the HypnoS probe. a, Structural composition of the probe. b, One-photon spectral properties. c, Binding affinity analysis, with HypnoS-mut as a control. d, Sub-second kinetic response. e, High specificity. f, Negligible enzymatic activity of the probe.
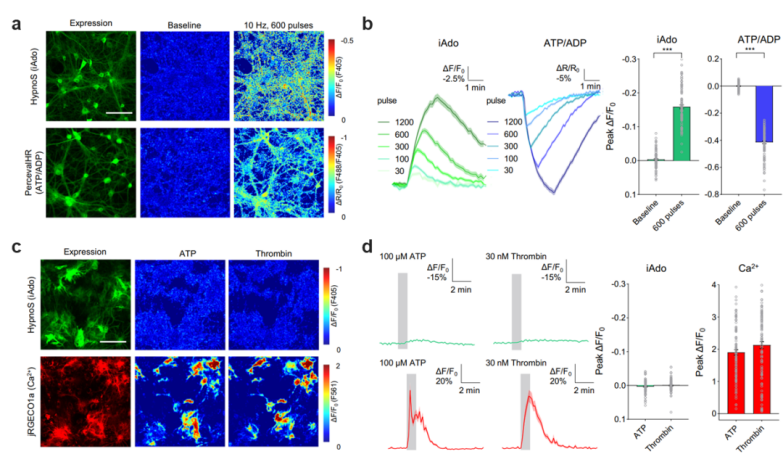
At the cellular level, the differences in intracellular adenosine (iAdo) generation, metabolism, and release among various brain cell types remain unclear, and there is ongoing debate over whether extracellular adenosine (eAdo) primarily originates from neurons or neighboring glial cells. Leveraging the genetically encodable nature of the novel HypnoS probe, the authors achieved cell-type-specific monitoring of iAdo dynamics by selectively expressing HypnoS in neurons and astrocytes across multiple systems, including primary cultured cells, acute brain slices, and freely moving mice. The study revealed marked differences in the levels, kinetics, and release patterns of adenosine between neurons and astrocytes (Figure 2), suggesting distinct contributions of these two cell types to adenosine homeostasis.
Figure 2. Measuring adenosine and ATP metabolism differences in primary neurons and astrocytes. a–b, Relationship between neuronal activity, adenosine metabolism, and ATP energy usage. c–d, Intracellular adenosine dynamics during calcium activity in astrocytes.
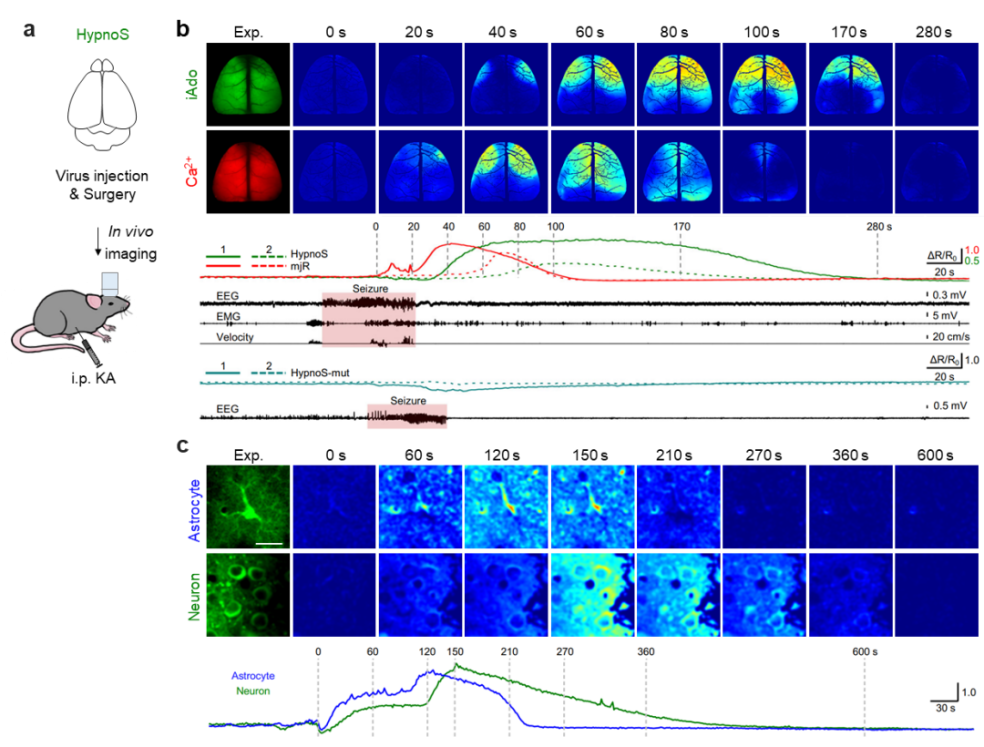
In pathological conditions such as epilepsy, adenosine is considered a key endogenous anticonvulsant factor. By combining the HypnoS probe with wide-field in vivo imaging, the authors visualized, for the first time, iAdo dynamics across the entire mouse cortex during acute seizures. They found that iAdo changes lagged significantly behind seizure-induced neuronal calcium activity and persisted for a longer duration—consistent with the proposed neuroprotective role of iAdo following seizures (Figure 3). Furthermore, using two-photon in vivo imaging at the single-cell level, the study revealed that astrocytes degrade iAdo more rapidly than neurons, expanding the understanding of cell-type-specific adenosine regulation during epileptic episodes.
Figure 3. Tracking intracellular adenosine dynamics during seizures with HypnoS. a, Schematic overview. b, Spatiotemporal relationship between HypnoS and calcium signals during seizures via wide-field imaging. c, Neuron vs. astrocyte iAdo dynamics during seizures revealed by two-photon in vivo imaging.
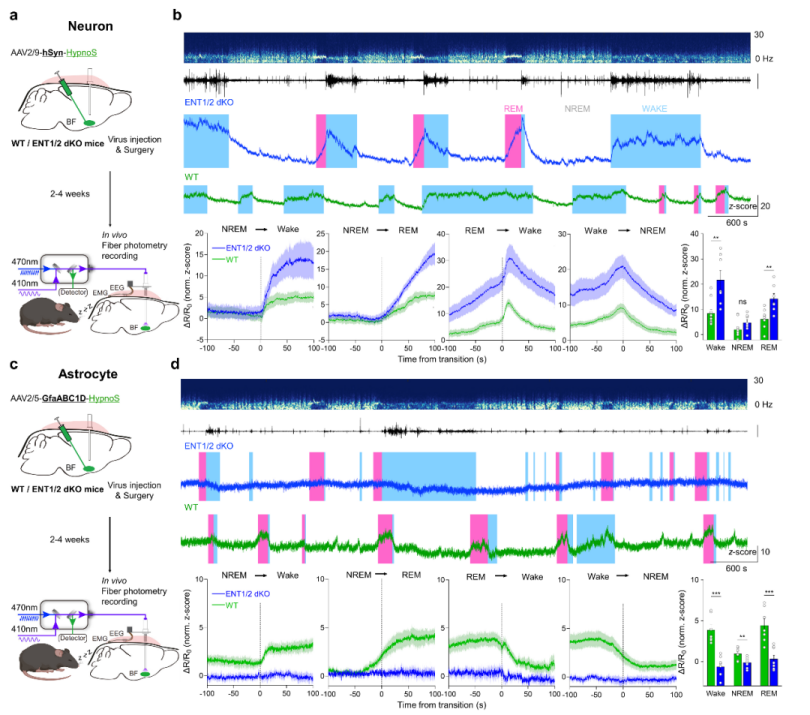
Under physiological conditions such as the sleep–wake cycle, adenosine is recognized as a critical modulator of sleep homeostasis. The research team used HypnoS in combination with fiber photometry, EEG, and EMG recordings in the mouse basal forebrain, capturing intracellular adenosine elevations during wakefulness and REM sleep, and declines during NREM sleep. The study demonstrated that ENT1/2 transporters mediate adenosine release in neurons but facilitate uptake in astrocytes, revealing a division of labor between cell types in maintaining sleep homeostasis (Figure 4). This finding provides a molecular foundation for targeting adenosine signaling in sleep disorder interventions.
Figure 4. Intracellular adenosine dynamics and cell-specific regulation during the sleep–wake cycle. a–b, Role of ENT1/2-mediated adenosine transport in neurons. c–d, Role of ENT1/2-mediated adenosine transport in astrocytes.
For those interested in the adenosine sensor described in the original article, please email the corresponding author and the first author! Original article link:https://www.nature.com/articles/s41467-025-59530-7