Literature Review | Nature | Adenosine Signaling in Astrocytes Coordinates Brain Metabolism and Function
Release time:2025-05-15 09:26:17
In the brain, neurons lack substantial metabolic reserves and therefore require a continuous supply of energy substrates. Astrocytes, in contrast, are capable of storing chemical energy in the form of glycogen and can respond to increased neuronal activity by rapidly activating glucose metabolism. Existing evidence suggests that the metabolic coupling between neurons and astrocytes plays a critical role in maintaining the proper function of neural circuits that govern core behaviors.
However, it remains unclear how astrocytes precisely sense the metabolic demands of nearby neurons, and which extracellular and intracellular signaling pathways regulate astrocytic glucose metabolism to support normal neuronal activity.
A study published in Nature on July 3, 2024, titled “Adenosine signalling to astrocytes coordinates brain metabolism and function,” sheds light on this mechanism. It reveals that metabolic activation of astrocytes, which is essential for neuronal activity, is mediated by the neuromodulator adenosine acting on A2B receptors expressed on astrocytes. Upon stimulation of these receptors, the cyclic AMP (cAMP)–protein kinase A (PKA) signaling pathway is activated, rapidly triggering glucose metabolism in astrocytes. This, in turn, leads to the release of lactate, which helps sustain fundamental brain functions such as sleep and memory.
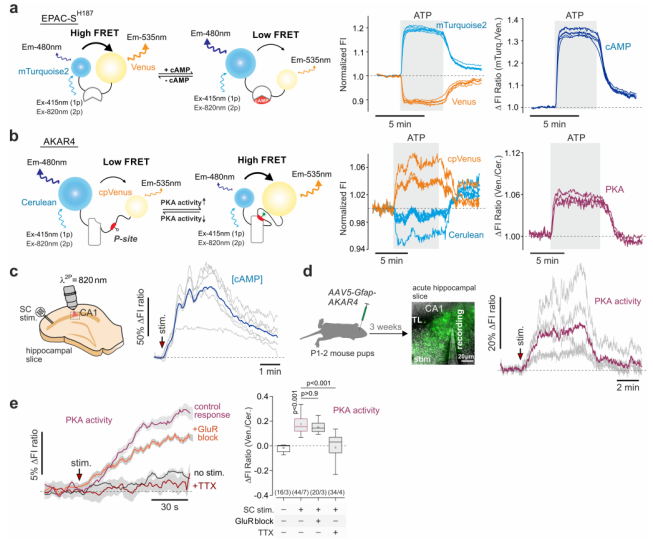
Researchers employed genetically encoded fluorescent sensors—Epac-SH187 for cAMP and AKAR4 for PKA activity—to investigate signaling dynamics in astrocytes. These sensors were expressed under the control of the GFAP promoter, allowing for real-time monitoring of changes within astrocytes in the CA1 region of the rat hippocampus (Fig. 1a-b). The results showed that stimulation of the Schaffer collateral (SC) fibers significantly elevated both cAMP levels and PKA activity in astrocytes. Interestingly, this elevation was not suppressed when glutamate receptor blockers (GluR block) were applied, suggesting that the neuronal activation of the cAMP–PKA signaling pathway in astrocytes occurs via signals other than glutamate (Fig. 1c-e, Fig. 2a-b).
Figure 1. Recordings of cAMP and PKA activity changes in astrocytes.
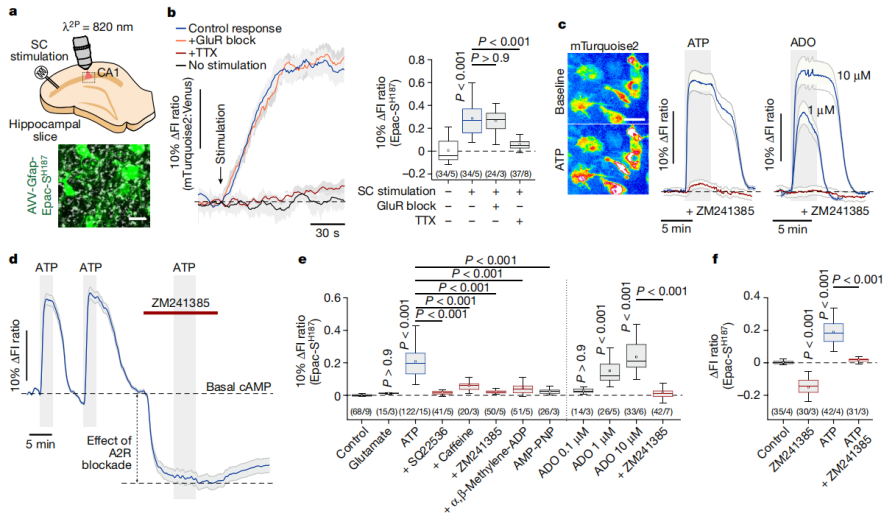
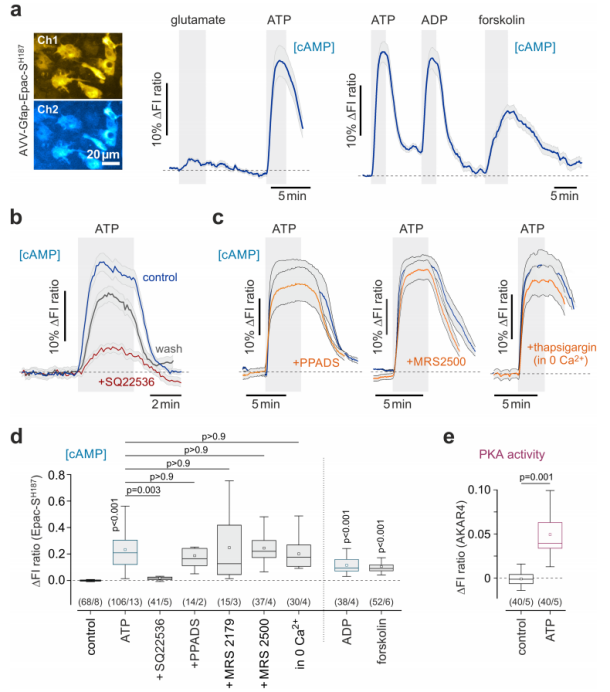
Numerous studies have indicated that increased neuronal activity promotes purine release into the extracellular space. The researchers found that purine nucleotides ATP and ADP elevated intracellular cAMP levels and PKA activity in astrocytes within brain slices (Fig. 2c, Fig. 3a). The adenylyl cyclase activator forskolin significantly boosted astrocytic cAMP levels, whereas the adenylyl cyclase inhibitor SQ22536 markedly suppressed the cAMP response induced by ATP. In contrast, blocking ionotropic purinergic receptors P2X (using PPADS), metabotropic purinergic receptors P2Y (using MRS 2179 and MRS 2500), or inhibiting calcium activity (via thapsigargin and calcium-free conditions) had no effect on ATP-induced changes in cAMP activity (Fig. 3b-e).
Furthermore, adenosine (ADO) was also found to strongly increase cAMP levels and PKA activity in astrocytes, an effect that was completely abolished by the adenosine A2 receptor antagonist ZM241385 (Fig. 2c-f). Collectively, these findings indicate that ATP’s influence on the astrocytic cAMP–PKA pathway is indirect and independent of intracellular calcium signaling. Instead, it is mediated by adenosine generated from extracellular ATP through the enzymatic activity of ectonucleotidases.
Figure 2. Neuronal activity recruits the cAMP–PKA signaling pathway in astrocytes.
Figure 3. ATP-induced activation of the cAMP signaling pathway in astrocytes.
2. Adenosine A2B Receptors in Astrocytes
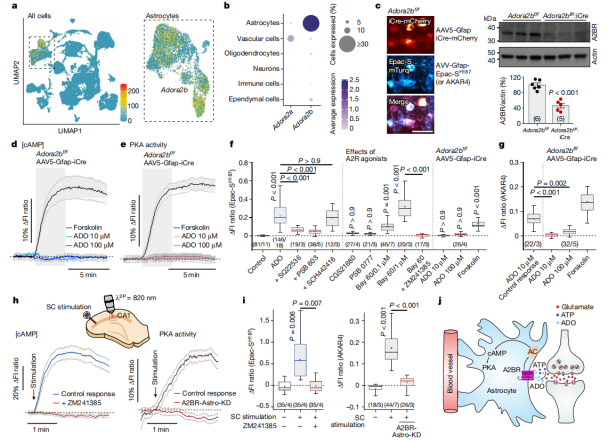
Single-cell RNA sequencing (RNA-Seq) analysis of the mouse brain revealed that adenosine A2B receptors (Adora2b) are highly and specifically expressed in astrocytes, while adenosine A2A receptors (Adora2a) show little to no expression in these cells (Fig. 4a-b). Experimental data showed that the A2B receptor antagonist PSB 603 effectively blocked the cAMP response induced by ADO in astrocytes, whereas the A2A receptor antagonist SCH442416 had no effect. Similarly, the A2B receptor agonist BAY 60 could mimic the effect of ADO on intracellular cAMP, while the A2A receptor agonist CGS21680 could not (Fig. 4f).
In Adora2b^f/f mice, A2B receptors were selectively deleted in astrocytes by delivering iCre recombinase via the AAV5-Gfap-iCre-mCherry vector. Following this deletion, even treatment with high concentrations of ADO (up to 100 μM) failed to induce any increase in cAMP or PKA activity. However, the cAMP and PKA responses triggered directly by forskolin remained unaffected (Fig. 4c-g). Additionally, blocking A2 receptors using ZM241385 or genetically deleting A2B receptors (creating the A2BR-Astro-KD model) both abolished the SC stimulation-induced increase in cAMP and PKA activity within astrocytes in the CA1 region (Fig. 4h-j).
These findings clearly demonstrate that adenosine mediates the neuron activity-dependent recruitment of the cAMP–PKA signaling pathway in astrocytes specifically through activation of the A2B receptor.
Figure 4. Neuron Activity-Dependent Recruitment of the cAMP–PKA Pathway in Astrocytes Is Mediated by Adenosine A2B Receptors
3. Adenosine Signaling and Brain Metabolism
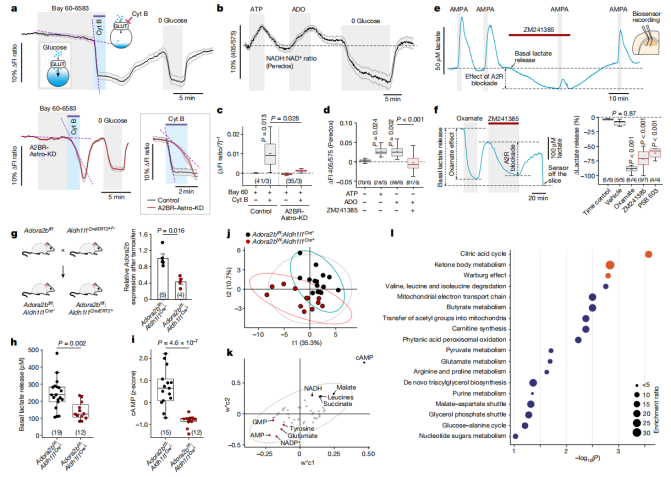
Using genetically encoded fluorescent sensors—FLIP12glu-700μΔ6 for glucose and Peredox for monitoring cytosolic NADH/NAD⁺ redox states—researchers observed that both ATP and ADO significantly enhanced the glucose consumption and glycolytic rates in astrocytes (Fig. 5a-d). Similar effects were observed with the A2B receptor agonist BAY 60-6583, but these effects were abolished in astrocytes lacking A2B receptors (A2BR-Astro-KD group) (Fig. 5a).
Next, enzyme-based microelectrode biosensors were used to monitor lactate levels in acute rat brain slices. The results showed that both ATP and ADO triggered substantial lactate release. Blocking A2 receptors with ZM241385 significantly reduced AMPA-induced lactate release. Inhibition of lactate dehydrogenase with Oxamate reduced basal lactate release by approximately 90%, while blocking A2B receptors with either PSB 603 or ZM241385 reduced lactate release by 60–70% (Fig. 3e-f).
To further validate these findings, Adora2b^f/f mice were crossed with Aldh1l1Cre/ERT2^+/- mice to generate a conditional A2B receptor knockout model specifically in astrocytes. After tamoxifen treatment, A2B receptor transcript levels in the brain were reduced by 60%, basal lactate release from brain slices decreased by 41%, and cAMP levels were also significantly lowered (Fig. 5g-i).
Targeted metabolomic analysis of brain tissue revealed that loss of A2B receptor function led to substantial metabolic reprogramming. Processes such as the tricarboxylic acid (TCA) cycle, ketone body metabolism, and the Warburg effect were significantly downregulated (Fig. 5j-l). These results highlight the essential role of A2B receptor-mediated signaling in regulating astrocytic glucose metabolism and sustaining overall brain metabolic activity.
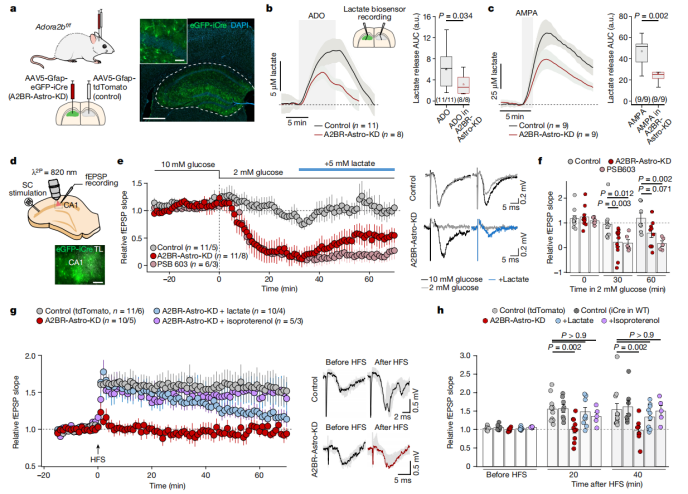
The hippocampal neural circuit plays a critical role in learning and memory. In experiments using Adora2b^f/f mice, researchers injected either AAV5-Gfap-eGFP-iCre (to create the A2BR-Astro-KD group) or AAV5-Gfap-tdTomato (as Control) into the hippocampus. The results showed that lactate release induced by either ADO or AMPA was reduced in the A2BR-Astro-KD group (Fig. 6a-c).
In acute hippocampal slice experiments, researchers evaluated synaptic function by recording Schaffer collateral field excitatory postsynaptic potentials (fEPSPs) while reducing the glucose concentration in the culture medium from 10 mM to 2 mM. In wild-type mouse slices, synaptic activity was maintained under low glucose conditions. However, in slices from mice with A2B receptor knockdown or pharmacological inhibition, synaptic activity was lost within 30 minutes. Remarkably, supplementation with lactate partially restored excitatory synaptic transmission (Fig. 6d-f).
Furthermore, in hippocampal slices, classical high-frequency stimulation (HFS) was used to induce long-term potentiation (LTP). The results showed that A2B receptor knockout mice failed to generate LTP. However, this impairment could be partially rescued by lactate supplementation or by applying the β-adrenergic receptor agonist isoproterenol (Fig. 6g-h).
This series of experiments provides strong evidence that under conditions of high energy demand or limited energy supply, adenosine-mediated metabolic signaling through astrocytic A2B receptors is essential for maintaining synaptic activity and plasticity.
Figure 6. Adenosine A2B Receptor-Mediated Signaling in Astrocytes Supports Synaptic Activity and Plasticity
5. Regulation of Learning, Memory, and Sleep by Astrocytic Adenosine A2B Receptors
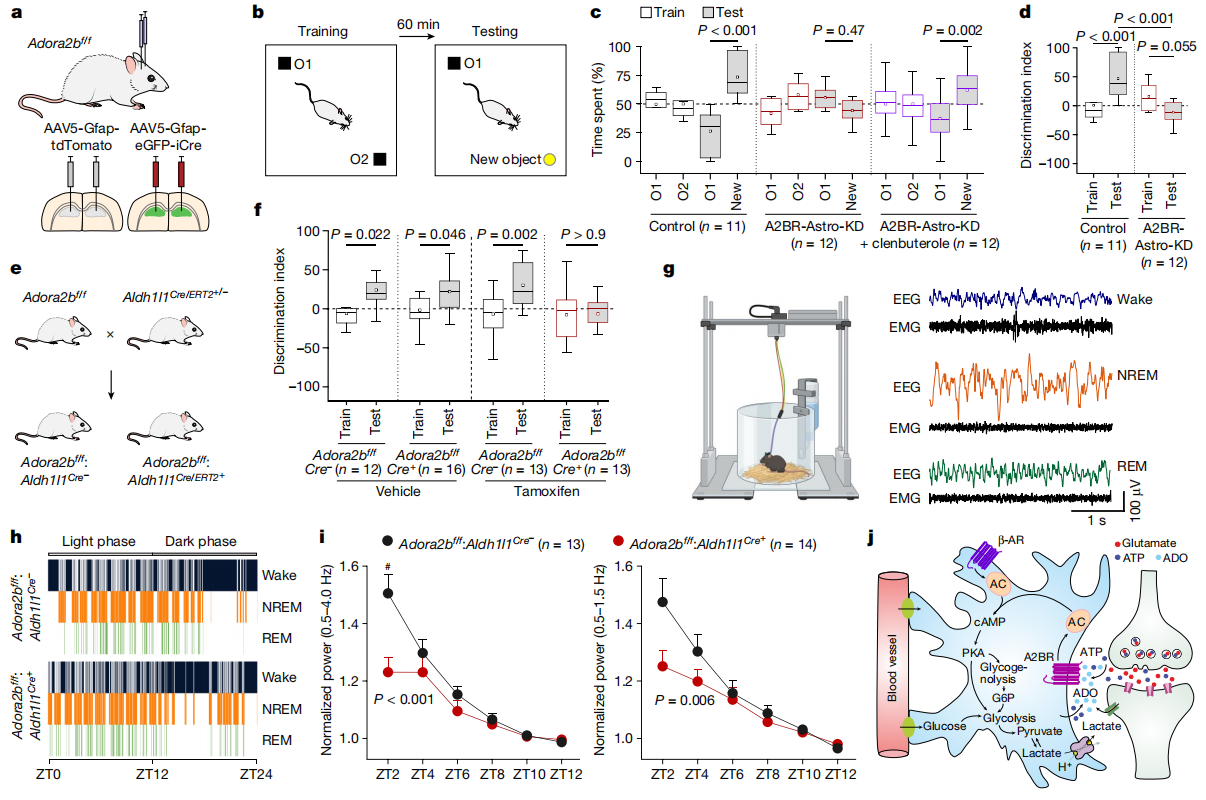
Researchers next evaluated the learning and memory abilities of mice using the novel object recognition (NOR) test. Control group mice spent significantly more time exploring the novel object compared to the familiar one, indicating intact recognition memory. In contrast, A2BR-Astro-KD mice showed no significant preference between the familiar and novel objects, suggesting impaired recognition memory (Fig. 7a–d). Similarly, conditional knockout mice (Adora2b^f/f : Aldh1l1Cre/ERT2^+/-) also exhibited severe deficits in recognition memory (Fig. 7e–f). Treatment with the β-adrenergic receptor agonist clenbuterol (0.25 mg/kg; intraperitoneal injection) partially restored recognition memory in A2B receptor knockdown mice (Fig. 7c).
Regarding sleep-wake regulation, specific deletion of A2B receptors in brain astrocytes led to fragmented sleep and wake states during the light phase (resting period), reduced non-rapid eye movement (NREM) sleep, and increased frequency of awakenings (Fig. 7g–h). To assess sleep pressure, researchers analyzed slow-wave activity during NREM sleep. A2B receptor-deficient mice exhibited significantly reduced overall slow-wave activity (0.5–4.0 Hz) and low-frequency slow waves (0.5–1.5 Hz), indicating decreased sleep pressure (Fig. 7i–j).
These findings demonstrate that adenosine A2B receptor-mediated signaling in astrocytes plays a crucial role in regulating learning and memory as well as maintaining consolidated sleep architecture and promoting synchronized slow-wave activity during NREM sleep.
In summary, this study identifies the adenosine A2B receptor as a critical astrocytic sensor of neuronal activity. It demonstrates that the cAMP–PKA signaling pathway in astrocytes regulates brain energy metabolism, thereby supporting fundamental brain functions such as sleep and memory. These findings provide key insights into the mechanisms that coordinate brain metabolism and function, and they offer potential targets and theoretical foundations for the treatment of related neurological disorders.