Customer Article | Nature Communications | Teams led by Zhi Zhang, Xia Zhu, and Wei Wang at USTC reveal how the nose–brain axis regulates behavioral responses to rancid odors
Release time:2026-01-20 13:50:27
The odor of spoiled food can trigger defensive reactions in humans—such as nausea, vomiting, and aversion—thereby helping to prevent metabolic disorders, yet the underlying neural mechanisms remain unclear. Traditional animal models for vomiting research (e.g., cats and dogs) lack robust molecular and cellular experimental tools. Although mice do not vomit, they exhibit retching-like behaviors, making them suitable for mechanistic studies. How are spoiled food odors transmitted through the olfactory system to the brain, and how do they subsequently regulate retching-like behaviors and aversive responses?
A collaborative study led by Professor Zhi Zhang and Professor Xia Zhu at the University of Science and Technology of China (USTC), together with Professor Wei Wang from the Department of Endocrinology at the First Affiliated Hospital of USTC, and Associate Researcher Qixin Zhou from the Kunming Institute of Zoology, Chinese Academy of Sciences, was published in Nature Communications under the title “A nose-to-brain axis for spoiled food odor-triggered defensive responses in male mice.”
This study demonstrates that 2-methylbutyric acid (2MBA) triggers defensive responses in male mice via two independent “nose–brain axis” neural pathways. One pathway—nose → olfactory bulb (OB) → anterior piriform cortex (aPir) → mediodorsal thalamic nucleus (MD) → ventrolateral periaqueductal gray (VLPAG) → ventral respiratory group (VRG) → respiratory muscles—mediates retching-like behavior. The other pathway—nose → OB → aPir → nucleus accumbens (NAc)—mediates aversive responses. These two pathways are implemented through projections from glutamatergic neurons in the aPir (aPir^Glu) to glutamatergic neurons in the MD (MD^Glu) and to GABAergic neurons in the NAc (NAcGABA), respectively.
2MBA induces retching-like behavior and aversive responses in male mice
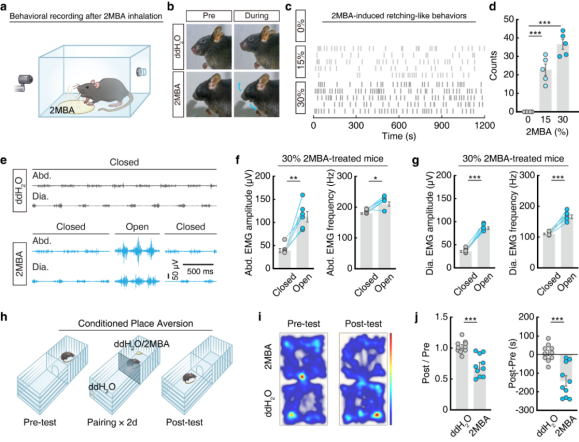
Inspired by the defensive reactions of humans to unpleasant odors, the authors established a paradigm to induce odor-evoked retching-like behavior in male mice. Filter papers containing aversive odorants at different concentrations (diluted in double-distilled water), including 2MBA, fenchone (FEN), and hexanol (HEX), were placed in a chamber to allow volatilization. Mice were allowed to freely inhale the odors for 20 minutes, while their behaviors were recorded by video (Fig. 1a). Among the tested odorants, only 2MBA (a rancid odor associated with spoiled food) elicited human-like retching mouth-opening movements in mice, and the frequency of these movements increased in a concentration-dependent manner (Fig. 1b–d).
A key physiological hallmark of retching is the activation of the diaphragm and abdominal muscles. Electromyography (EMG) recordings revealed that during 2MBA-induced mouth-opening movements, mice exhibited significantly increased amplitude and frequency of activity in the diaphragm and external oblique abdominal muscles. Such changes were not observed in mice exposed to other odors or in control conditions (Fig. 1e–g). Accordingly, this phenotype was defined as retching-like behavior.
To further assess aversive responses, a conditioned place aversion (CPA) test was performed (Fig. 1h). Mice in the 2MBA-paired group spent less time in the corresponding chamber and showed higher CPA scores (Fig. 1i–j), confirming that 2MBA induces aversion in mice. After establishing an anosmia model by intranasal irrigation with zinc sulfate, mice exposed to 2MBA no longer exhibited retching-like behavior or aversive responses, whereas sham-operated mice showed robust responses. These results indicate that the defensive responses triggered by 2MBA are dependent on olfactory perception.
Figure 1 | 2MBA-induced retching-like behavior and aversive responses in mice
2MBA activates aPirGlu neurons to trigger retching-like behavior and aversive responses
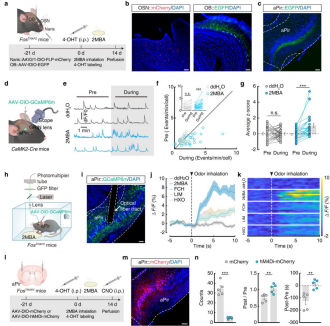
It is well established that olfactory sensory neurons (OSNs) in the olfactory epithelium project to the OB. To identify downstream brain regions activated by 2MBA, the authors used FosTRAP2 mice. Neurons were specifically labeled by injecting AAVs into the nasal cavity and OB, followed by intraperitoneal injection of 4-hydroxytamoxifen (4-OHT), after which mice were exposed to 2MBA (Fig. 2a). The results showed that EGFP⁺ fibers downstream of OB neurons were highly enriched in the aPir, whereas other olfactory cortical regions—such as the posterior piriform cortex (pPir), olfactory tubercle (OT), and cortical amygdala (CoA)—showed minimal labeling (Fig. 2b–c). Accordingly, subsequent analyses focused on the role of the aPir.
By crossing FosTRAP2 mice with Ai14-tdTomato reporter mice and administering 4-OHT to label Fos⁺ activated neurons, the authors confirmed that glutamatergic neurons in the aPir (aPirGlu) respond robustly to 2MBA.
Using miniscope imaging in combination with the calcium indicator GCaMP6m (Fig. 2d), they found that approximately 30% of aPir neurons were selectively activated by 2MBA alone, while 33.3% were co-activated by 2MBA and other odors (Fig. 2e–g). Fiber photometry recordings further showed that 2MBA induced significantly higher calcium signals (ΔF/F) in aPirGlu neurons compared with other odors, and that these responses were concentration dependent (Fig. 2h–k), demonstrating selective responsiveness of aPirGlu neurons to 2MBA.
Chemogenetic inhibition of 2MBA-activated aPirGlu neurons (via injection of AAV-DIO-hM4Di-mCherry and CNO administration; Fig. 2l–m) markedly reduced both the frequency of retching-like behaviors and CPA scores in mice (Fig. 2n). c-Fos immunostaining confirmed a TRAP labeling efficiency of 92.35%, indicating that enhanced activity of aPirGlu neurons is a critical neural substrate underlying 2MBA-induced defensive responses.
Figure 2 | 2MBA activates aPirGlu neuronal activity to trigger retching-like behavior and aversive responses
2MBA-activated aPirGlu neurons project to the mediodorsal thalamus (MD)
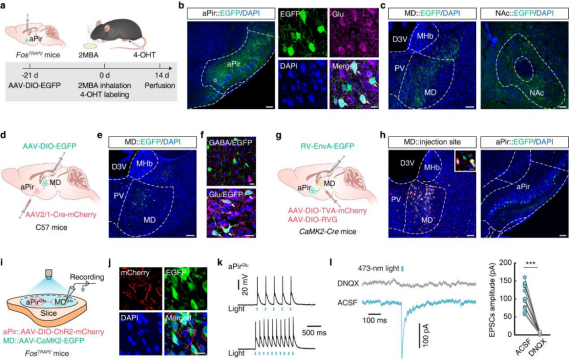
To identify the downstream targets of aPirGlu neurons involved in 2MBA-induced defensive responses, AAV-DIO-EGFP was injected into the aPir of FosTRAP2 mice. Three weeks later, mice received intraperitoneal injection of 4-hydroxytamoxifen (4-OHT) and were exposed to 2MBA (Fig. 3a). Two weeks thereafter, EGFP⁺ neurons were observed in the aPir (Fig. 3b), and EGFP⁺ fibers were detected in the MD, NAc, lateral hypothalamus (LH), and paratenial nucleus (PT) (Fig. 3c).
Activity mapping in FosTRAP2:Ai14 mice revealed that following 2MBA exposure, tdTomato expression was significantly increased in the MD—where 84% of labeled neurons colocalized with glutamatergic markers—and in the NAc, where 93% colocalized with GABAergic markers, whereas no significant changes were observed in the LH or PT. Accordingly, subsequent analyses focused on the aPirGlu→MD and aPirGlu→NAc pathways.
Given the established role of the MD in sensory information processing, the authors first examined the aPirGlu→MD pathway using anterograde monosynaptic tracing. AAV2/1-Cre-mCherry was injected into the aPir, and AAV-DIO-EGFP was injected into the MD (Fig. 3d). Three weeks later, EGFP⁺ neurons in the MD exclusively colocalized with glutamatergic markers (Fig. 3e,f), confirming that aPir neurons project to glutamatergic MD neurons (MDGlu).
To further define the circuit architecture, retrograde monosynaptic tracing was performed by injecting helper viruses into the MD of CaMK2-Cre mice, followed three weeks later by injection of EnvA-pseudotyped RV-ΔG-EGFP (Fig. 3g). One week later, EGFP⁺ neurons colocalizing with glutamatergic markers were detected in the aPir (Fig. 3h), confirming direct projections from aPirGlu to MDGlu neurons.
To validate functional connectivity, AAV-DIO-ChR2-mCherry was injected into the aPir and AAV-CaMK2-EGFP into the MD of FosTRAP2 mice (Fig. 3i). After 4-OHT administration and 2MBA exposure, mCherry⁺ fibers and EGFP⁺ neurons were observed in the MD (Fig. 3j). Optogenetic stimulation of aPirGlu terminals evoked excitatory postsynaptic currents (EPSCs) in MDGlu neurons, which were abolished by the AMPA receptor antagonist DNQX (Fig. 3k,l), demonstrating a functional synaptic connection.
Figure 3 | 2MBA-activated aPirGlu neurons project to the MD
2MBA-activated aPirGlu neurons project to the nucleus accumbens (NAc)
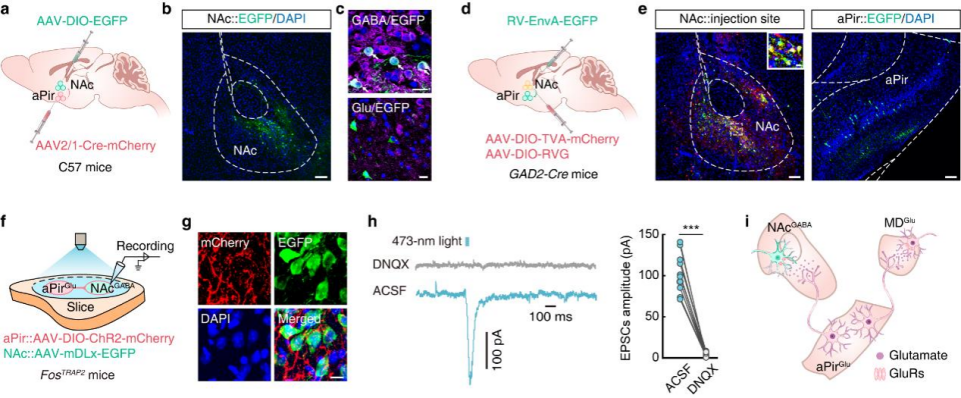
Previous studies have shown that the NAc—composed of approximately 95% GABAergic neurons—plays an important role in odor processing. To verify the connectivity of the aPirGlu→NAc pathway, AAV2/1-Cre-mCherry was injected into the aPir and AAV-DIO-EGFP into the NAc of C57 mice (Fig. 4a). Three weeks later, EGFP⁺ neurons in the NAc exclusively colocalized with GABAergic markers (Fig. 4b,c).
In GAD2-Cre mice, helper viruses were injected into the NAc followed by RV-ΔG-EGFP (Fig. 4d). One week later, EGFP⁺ neurons colocalizing with glutamatergic markers were detected in the aPir (Fig. 4e), confirming that aPirGlu neurons project to GABAergic neurons in the NAc (NAcGABA).
To assess functional connectivity, AAV-DIO-ChR2-mCherry was injected into the aPir and AAV-mDLx-EGFP into the NAc of FosTRAP2 mice (Fig. 4f). After 4-OHT treatment and 2MBA exposure, mCherry⁺ fibers and EGFP⁺ neurons were observed in the NAc (Fig. 4g). Optogenetic stimulation elicited EPSCs in NAcGABA neurons, which were abolished by DNQX (Fig. 4h), confirming a functional synaptic connection.
To determine whether aPir neurons projecting to the MD and NAc belong to the same population, retro-AAV-EGFP was injected into the MD and retro-AAV-mCherry into the NAc of C57 mice. The results showed that 75.44% of labeled neurons in the aPir did not overlap, indicating that distinct subpopulations of aPir neurons project to the MD and NAc, potentially mediating different behaviors. Collectively, these findings demonstrate that 2MBA-activated aPirGlu neurons project to both MD^Glu and NAcGABA neurons (Fig. 4i).
Figure 4 | 2MBA-activated aPirGlu neurons project to the NAc
The nose → OB → aPir → MD pathway is essential for 2MBA-induced retching-like behavior
To determine the functional roles of the aPirGlu→MDGlu and aPirGlu→NAcGABA pathways, in vivo multi-electrode electrophysiological recordings were performed in the MD and NAc of freely moving mice (Fig. 5a). During retching-like behavior, neuronal firing rates in the MD increased significantly, whereas no change was observed in the NAc (Fig. 5b,c). Conversely, during the post-test phase of the CPA paradigm, neuronal firing rates in the NAc increased when mice entered the 2MBA-paired chamber, while MD activity remained unchanged. These results suggest that the MD primarily regulates retching-like behavior, whereas the NAc mediates aversive responses.
Fiber photometry recordings in CaMK2-Cre mice expressing AAV-DIO-GCaMP6m in the MD revealed time-locked increases in MDGlu calcium signals during 2MBA-induced retching-like behavior, but not during CPA testing (Fig. 5d–f). Single-cell calcium imaging using GRIN lenses (Fig. 5g) further confirmed that MD neurons innervated by the aPir exhibited increased calcium activity exclusively during retching-like behavior and showed no response to other odors (Fig. 5h,i), indicating specificity for 2MBA-induced retching.
Using multisite injections of pathway-specific viruses combined with RV-Flp-DsRed tracing (Fig. 5j), EGFP⁺/DsRed⁺ co-labeled neurons were observed in the OB (Fig. 5k), enabling visualization of the complete nose → OB → aPir → MD pathway. Chemogenetic inhibition of this pathway via CNO administration to the aPir (Fig. 5l) significantly reduced retching-like behavior without affecting conditioned aversion (Fig. 5m–p), demonstrating that the nose → OB → aPir → MD pathway is necessary for 2MBA-induced retching-like behavior, but not aversion.
Figure 5 | The nose → OB → aPir → MD circuit regulates 2MBA-induced retching-like behavior
The nose → OB → aPir → NAc pathway is essential for 2MBA-induced aversive responses
To investigate the function of the aPirGlu→NAcGABA pathway, fiber photometry recordings were performed in GAD2-Cre mice expressing AAV-DIO-GCaMP6m in the NAc (Fig. 6a). Calcium activity in the NAc increased when mice entered the 2MBA-paired chamber during CPA post-testing, but showed no change during retching-like behavior (Fig. 6b,c).
Single-cell calcium imaging using GRIN lenses (Fig. 6d) further confirmed these findings (Fig. 6e–g). Notably, NAc neurons were also activated during aversive responses to other unpleasant odors, indicating a broader role of the NAc in odor-induced aversion.
Using multisite viral injections combined with RV-Flp-DsRed tracing (Fig. 6h), EGFP⁺/DsRed⁺ co-labeled neurons were observed in the OB, clearly delineating the anatomical connectivity of the nose → OB → aPir → NAc pathway (Fig. 6i). Chemogenetic inhibition of this nose–brain axis via CNO administration to the aPir (Fig. 6j) attenuated conditioned aversion to 2MBA without affecting retching-like behavior (Fig. 6k–n), confirming that this pathway is essential for 2MBA-induced aversive responses.
Figure 6 | The nose → OB → aPir → NAc circuit regulates 2MBA-induced aversive responses
The nose–brain–muscle axis underlies 2MBA-induced retching-like behavior
Given that the nose → OB → aPir → MD pathway mediates retching-like behavior and that such behavior is accompanied by activation of the diaphragm and abdominal muscles, the authors further investigated how the MD regulates respiratory muscles. PRV-RFP was injected into the external oblique abdominal muscle and PRV-EGFP into the diaphragm of C57 mice (Fig. 7a). Four days later, co-labeled neurons were observed in the paraventricular nucleus (PVN), periaqueductal gray (PAG), and VRG (Fig. 7b).
Injection of AAV-DIO-mCherry into the MD and retrograde EGFP into the VRG revealed EGFP⁺ neurons in the VLPAG surrounded by mCherry⁺ fibers (Fig. 7c–e), suggesting that the MD regulates respiratory muscles via an MD → VLPAG → VRG pathway.
To validate the functional role of the MD → VLPAG pathway, AAV-DIO-ChR2-mCherry was injected into the MD of CaMK2-Cre mice, and optical fibers were implanted in the VLPAG. Upon activation with 473 nm blue light, mice exhibited increased retching-like behavior, along with elevated frequency and amplitude of external oblique and diaphragmatic muscle activity, whereas no such changes were observed in control mice.
Collectively, these findings demonstrate that rancid odors trigger distinct defensive responses through two independent pathways: Retching-like behavior via the nose → OB → aPir → MD → VLPAG → VRG → respiratory muscles axis Aversive responses via the nose → OB → aPir → NAc pathway (Fig. 7f)
Figure 7 | Definition of the MD → VLPAG → VRG → respiratory muscle circuit
Conclusion
This study, for the first time, elucidates a dual “nose–brain axis” mechanism by which the odor of spoiled food triggers defensive responses. It identifies aPirGlu neurons as a central hub of this circuitry and reveals a neural coding strategy that enables specific integration of olfactory information and selective behavioral outputs. These findings provide potential targets for the development of odor-based therapies and intranasal neuromodulation strategies, with applications in maintaining metabolic homeostasis and treating metabolic disorders.
Research Tools Used in This Study
The viral vector tools used in this work are available from Brain Case Biotech, In addition, we offers a wide range of customized services.Please contact bd@ebraincase.com