Customer Article丨Cell Reports丨Yi-Zheng Wang’s team from the Academy of Military Medical Sciences reveals how VTA astrocytes reverse anxiety–depression mechanisms via the adenosine pathway
Release time:2025-09-16 17:25:00
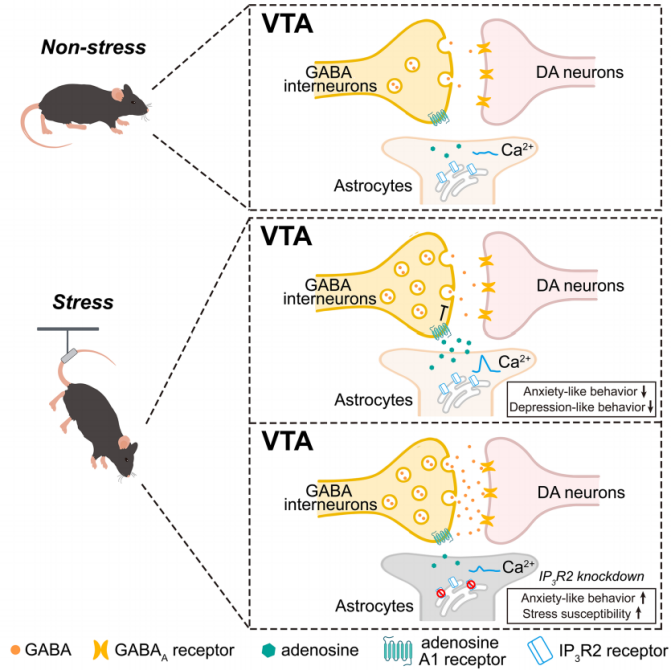
Anxiety and depression are major causes of disability. Depression patients often suffer from comorbid anxiety as well as dopaminergic dysfunction. Stress-induced abnormalities in ventral tegmental area (VTA) dopamine neuron function and dysregulation of mesolimbic dopamine release contribute to the onset of anxiety and depression. Studies have shown that VTA dopamine neurons play a key role in the pathophysiology of these disorders, but their direct pharmacological activation carries risks of addiction. Therefore, maintaining homeostatic activity of VTA dopamine neurons under stress is considered a safe and effective preventive strategy. Astrocytes, by releasing gliotransmitters or uptaking neurotransmitters, can modulate neuronal activity. Previous studies indicated that regulating VTA astrocytes could affect emotional behaviors by altering neuronal activity, but how VTA astrocytes regulate dopamine neurons during stress remained unclear.
On July 24, 2025, Yi-Zheng Wang’s team at the Academy of Military Medical Sciences published a research article in Cell Reports titled “Astrocytic modulation of GABA transmission in the VTA limits progression of anxiety- and depression-like behaviors in adult mice.” The study demonstrated that during stress responses, VTA astrocytes are activated via Ca²⁺ elevation and inhibit GABA transmission through adenosine A1 receptors on GABAergic interneurons. This disinhibits dopamine (DA) neurons, thereby alleviating anxiety- and depression-like behaviors. Disruption of VTA astrocytic Ca²⁺ signaling (e.g., by knocking down IP3R2) or inhibition of gliotransmitter release increased mice’s susceptibility to chronic stress and aggravated related behavioral disorders.
VTA astrocytes show specific responses to various stress stimuli
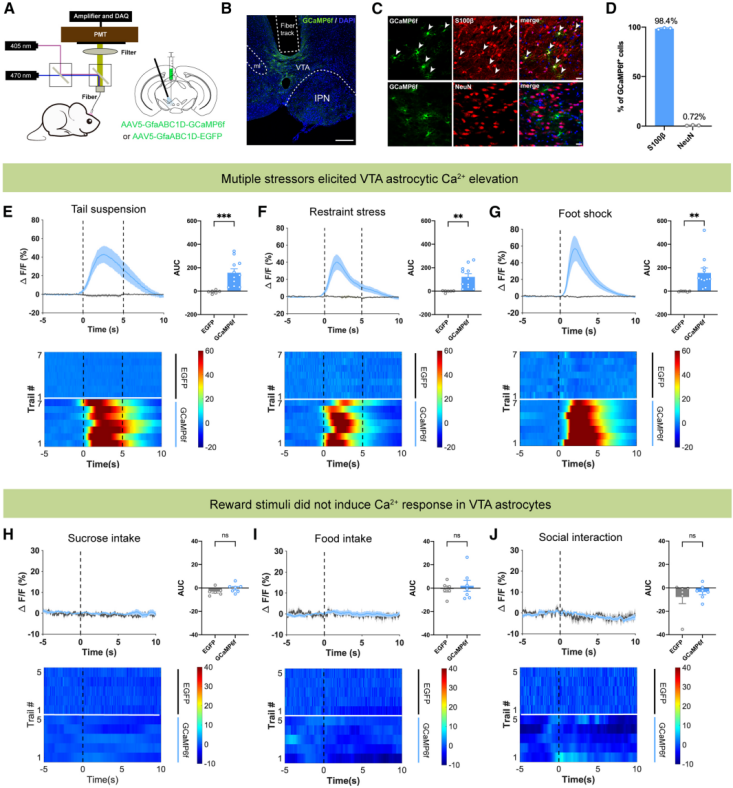
Previous studies have shown that astrocytic activity is associated with intracellular calcium (Ca²⁺) signaling. To investigate the potential role of VTA astrocytes in response to aversive stimuli, fiber photometry was used to monitor Ca²⁺ dynamics of VTA astrocytes in freely moving mice. An AAV5-GfaABC1D-GCaMP6f virus was injected into the VTA to specifically target astrocytes (Fig. 1A, 1B). Results showed that 98.4% of AAV-labeled cells were S100β⁺ astrocytes, while only 0.72% of GCaMP6f-expressing cells were neurons (Fig. 1C, 1D). Stress stimuli, such as tail suspension, restraint stress, and foot-shock stimulation, induced a significant increase in Ca²⁺ fluorescence signals in GCaMP6f-injected mice, with the AUC of Ca²⁺ transients markedly elevated during stress. In contrast, control mice showed no such responses (Fig. 1E–1G). Conversely, rewarding stimuli, such as sucrose/food consumption or social interaction with female mice, did not alter VTA astrocytic Ca²⁺ fluorescence in GCaMP6f-injected mice (Fig. 1H–1J).Taken together, these fiber photometry data indicate that VTA astrocytes are specifically activated by stress, but not by rewarding stimuli.
Figure 1. Stress-induced Ca²⁺ elevation in VTA astrocytes
Optogenetic activation of VTA astrocytes produces anxiolytic and antidepressant effects via dopamine neuron activation
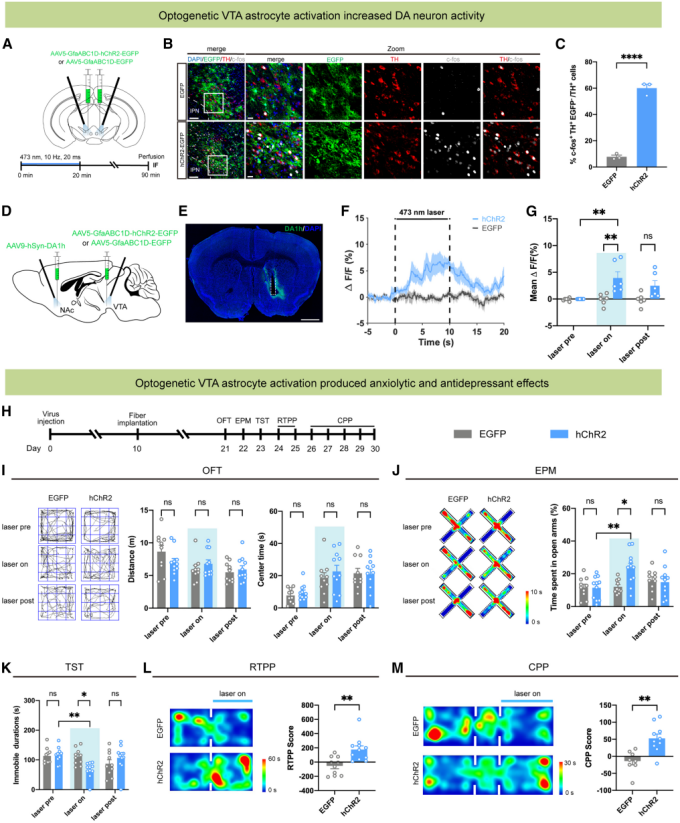
To investigate the impact of VTA astrocyte activation on dopamine neuron activity, AAV5-GfaABC1D-hChR2-EGFP was bilaterally injected into the VTA to specifically activate astrocytes. After 20 minutes of laser stimulation (473 nm, 10 Hz, 20 ms), the proportion of c-Fos⁺/TH⁺ double-labeled cells in the VTA of hChR2-injected mice was significantly higher than that of controls (Fig. 2A–2C). In the nucleus accumbens (NAc) expressing the dopamine sensor DA1h, dopamine fluorescence signals were markedly enhanced, suggesting increased dopamine release (Fig. 2D–2G).
Given that stress induces anxiety and depression, the authors further evaluated whether activating VTA astrocytes could produce anxiolytic and antidepressant effects in adult wild-type mice (Fig. 2H). In the open field test, no significant difference was observed between the hChR2 and control groups in total locomotor distance or time spent in the center (Fig. 2I). In the elevated plus maze, hChR2 mice spent a greater percentage of time in the open arms during laser-on sessions, while controls showed no such effect, indicating reduced anxiety-like behavior (Fig. 2J). In the tail suspension test, immobility time was reduced in hChR2 mice, suggesting an antidepressant effect (Fig. 2K).
Previous studies reported that antidepressant effects are often accompanied by positive valence states. Therefore, the authors examined whether optogenetic activation of VTA astrocytes could induce reward-related behaviors. In the real-time place preference (RTPP) test, hChR2 mice spent more time in the laser-paired chamber, with higher RTPP scores (Fig. 2L). In the conditioned place preference (CPP) test, hChR2-expressing mice developed a preference for the laser-paired chamber (Fig. 2M).
Taken together, activation of astrocytes in the ventral tegmental area enhances dopamine neuron activity, alleviates anxiety- and depression-like behaviors, and induces reward effects.
Figure 2. Optogenetic activation of VTA astrocytes enhances dopamine neuron activity and produces anxiolytic and antidepressant effects
Activation of VTA astrocytes enhances dopamine neuron activity via adenosine A1 receptors
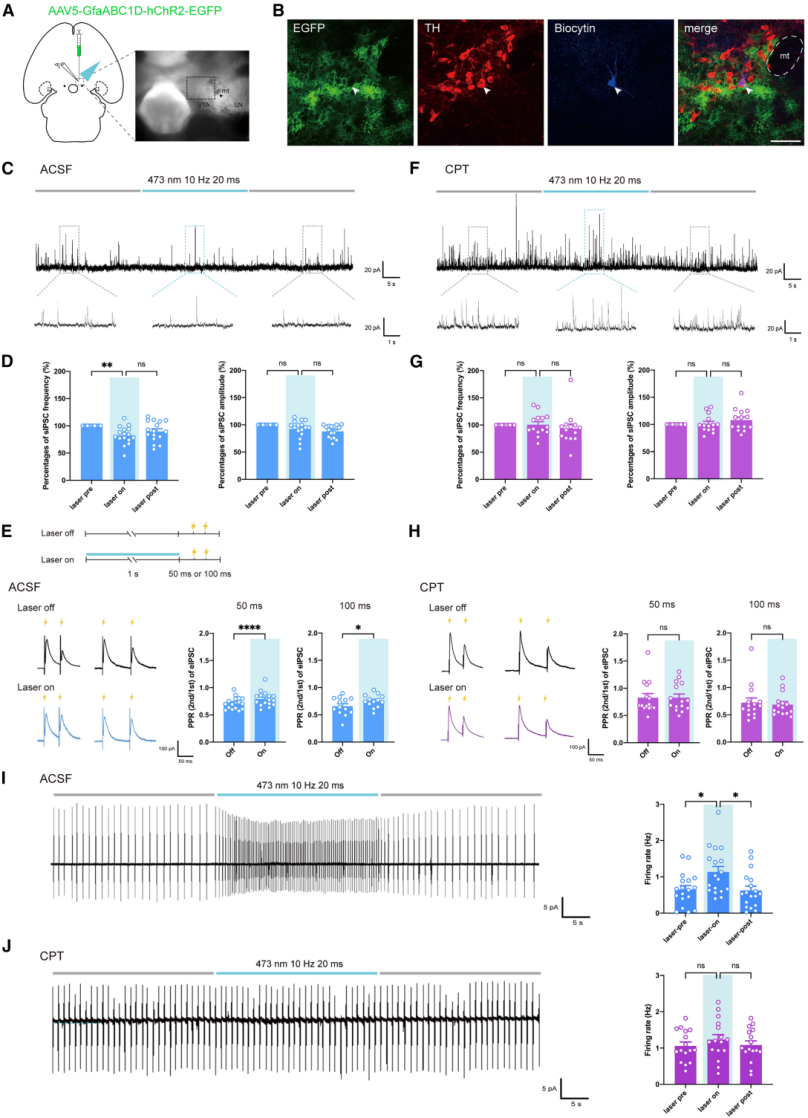
To explore how VTA astrocytes affect dopamine neuron activity, the study combined in vitro electrophysiological recordings of dopamine neurons with optogenetic activation of VTA astrocytes (Fig. 3A, 3B). In brain slices from hChR2-injected mice, optogenetic activation of VTA astrocytes reduced the frequency—but not the amplitude—of spontaneous inhibitory postsynaptic currents (sIPSCs) compared to pre-stimulation levels (Fig. 3C, 3D). The paired-pulse ratio (PPR) of inhibition was also increased (Fig. 3E). In slices from EGFP-injected mice, laser stimulation had no effect on sIPSCs or inhibitory PPR. These findings indicate that VTA astrocyte activation suppresses presynaptic GABA release, thereby relieving inhibition of dopamine neurons.
Given that ATP/adenosine serves as a major gliotransmitter and that adenosine kinase is highly expressed in VTA astrocytes, the authors hypothesized that synaptic GABA transmission is regulated through adenosine receptors. The reduction in sIPSC frequency and increase in inhibitory PPR induced by VTA astrocyte activation were abolished by the adenosine A1 receptor antagonist CPT (10 μM) (Fig. 3F–3H), suggesting that VTA astrocytes reduce inhibition of dopamine neurons via A1 receptors. Furthermore, optogenetic activation of VTA astrocytes increased the firing rate of dopamine neurons compared to baseline, an effect attenuated by CPT (Fig. 3I, 3J). In summary, activation of VTA astrocytes enhances dopamine neuron activity in an adenosine A1 receptor–dependent manner.
Activation of VTA astrocytes relieves inhibition of dopamine neurons via adenosine A1 receptors on GABAergic interneurons
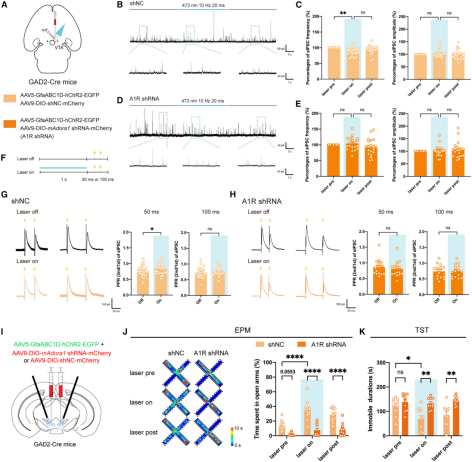
Since GABAergic interneurons in the VTA provide significant inhibitory input to dopamine neurons, it was hypothesized that VTA astrocytes may relieve dopamine neuron inhibition through presynaptic adenosine A1 receptors on GABAergic interneurons. To test this, AAV5-GfaABC1D-hChR2-EGFP was expressed in adult GAD2-Cre mice, followed by VTA injection of AAV9-EF1a-DIO-mAdora1 shRNA-mCherry to selectively knock down adenosine A1 receptor expression in VTA GABAergic interneurons (Fig. 4A). A shNC injection served as control.
Behavioral tests showed that, compared with controls, mice in the A1R shRNA group displayed reduced total locomotor distance and central time in the open field test, as well as decreased percentage of time spent exploring open arms in the elevated plus maze, indicating that A1R knockdown in VTA GABAergic neurons induced anxiety-like behaviors. No differences were observed between groups in sucrose preference, tail suspension, or forced swim tests.
Electrophysiological recordings revealed that, compared with controls, the effects of VTA astrocyte activation on sIPSC frequency and inhibitory PPR were abolished in A1R shRNA mice (Fig. 4A–4H), demonstrating that adenosine A1 receptors on VTA GABAergic interneurons are essential for astrocyte-induced disinhibition of dopamine neurons. Moreover, in behavioral assays, control mice exhibited increased open-arm time in the elevated plus maze and reduced immobility in the tail suspension test during laser stimulation, whereas these anxiolytic and antidepressant effects were absent in A1R shRNA mice (Fig. 4I–4K). Together, these results confirm that VTA astrocyte activation relieves inhibition of dopamine neurons through presynaptic adenosine A1 receptors on VTA GABAergic interneurons.
Figure 4. Knockdown of adenosine A1 receptors in VTA GABAergic neurons abolishes the effects of optogenetic activation of VTA astrocytes
Downregulation of IP3R2 in VTA astrocytes enhances inhibition of dopamine neurons
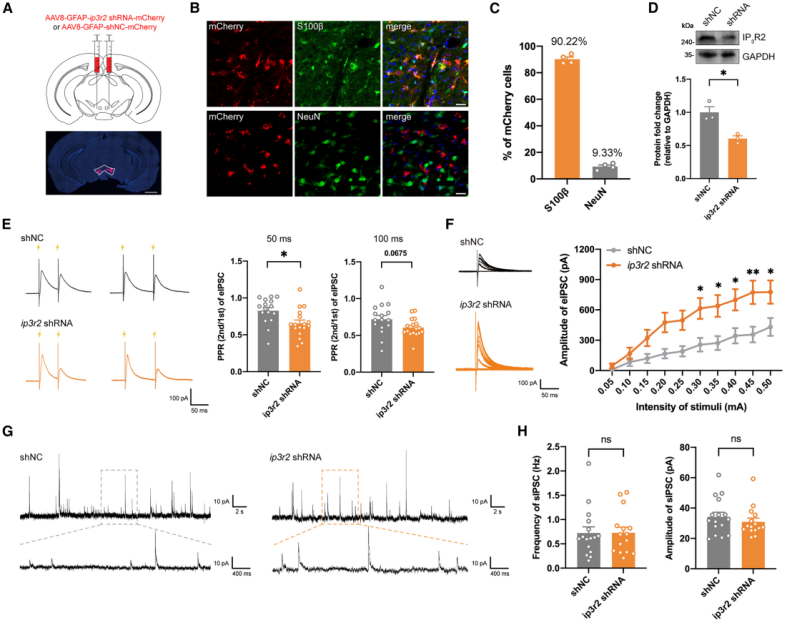
The above results indicate that enhanced Ca²⁺ dynamics in VTA astrocytes can increase dopamine neuron activity through a presynaptic adenosine A1 receptor–dependent mechanism. To disrupt Ca²⁺ dynamics in VTA astrocytes, the researchers expressed GFAP promoter–driven IP3R2 shRNA (AAV8-GFAP-ip3r2 shRNA-mCherry) in adult mice (Fig. 5A–5D). This manipulation weakened the Ca²⁺ elevation of astrocytes in response to aversive stimuli.
Downregulation of IP3R2 significantly reduced inhibitory PPR at a 50 ms interval and showed a decreasing trend at 100 ms (Fig. 5E). At stimulation intensities of 0.3–0.5 mA, the amplitude of evoked inhibitory postsynaptic currents (eIPSCs) in dopamine neurons was higher in the shRNA group than in controls (Fig. 5F), whereas the frequency and amplitude of sIPSCs showed no differences between groups (Fig. 5G, 5H). Together, these results suggest that knockdown of IP3R2 in VTA astrocytes enhances inhibition of dopamine neurons. Using an adenosine sensor to detect astrocyte-derived adenosine release in the VTA, the study further showed that in shNC mice, exposure to aversive stimuli induced a significant increase in adenosine fluorescence signals. In contrast, this increase was abolished in ip3r2 shRNA mice. These findings indicate that stress stimulates VTA astrocytes to release Ca²⁺-dependent adenosine, which in turn relieves inhibition of dopamine neurons.
Figure 5. Impaired Ca²⁺ elevation in VTA astrocytes enhances inhibition of dopamine neurons
Impairment of VTA astrocyte function induces anxiety-like behavior and increases susceptibility to chronic stress
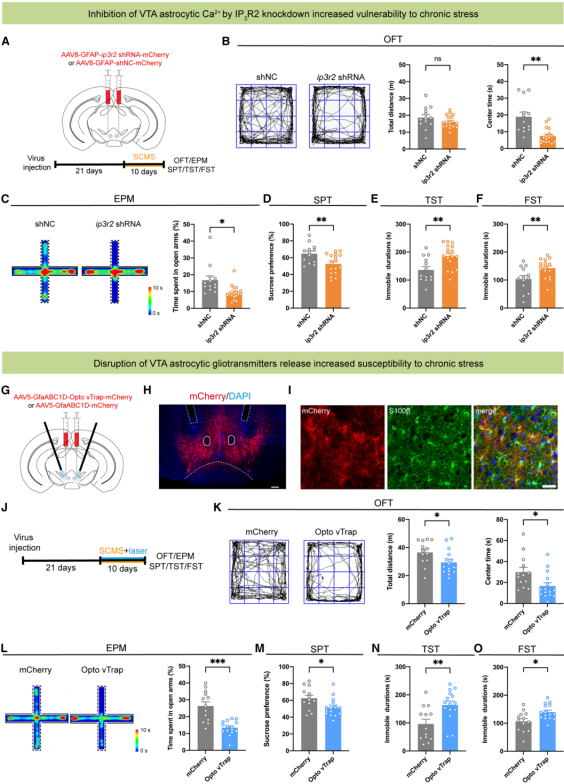
Subthreshold chronic mild stress (SCMS) was applied to mice to examine the effect of IP3R2 downregulation on chronic stress susceptibility (Fig. 6A). After 10 days of SCMS exposure, mice injected with ip3r2 shRNA exhibited anxiety-like phenotypes in the open field test and elevated plus maze (Fig. 6B, 6C), reduced sucrose preference (Fig. 6D), and increased immobility times in both the tail suspension test and the forced swim test (Fig. 6E, 6F).
To further test the role of gliotransmitter release, bilateral expression of AAV5-GfaABC1D-Opto vTrap-mCherry in the VTA was used to inhibit vesicular gliotransmitter release from astrocytes (Fig. 6G–6I). During laser stimulation, mice showed reduced percentage of time spent in open arms in the elevated plus maze, while no significant changes were observed in the open field or tail suspension tests. However, when combined with 10 days of SCMS exposure (Fig. 6J), Opto vTrap-mediated inhibition of gliotransmitter release caused a decrease in total locomotion and central area time in the open field test (Fig. 6K), reduced open-arm exploration in the elevated plus maze (Fig. 6L), further lowered sucrose preference, and prolonged immobility times in both the tail suspension and forced swim tests (Fig. 6M–6O).
Taken together, these results demonstrate that impaired Ca²⁺ elevation or blocked gliotransmitter release in VTA astrocytes induces anxiety-like behavior and increases susceptibility to chronic stress–induced depressive phenotypes, highlighting the protective role of VTA astrocytes in preventing the development of anxiety and depression.
Figure 6. Disruption of VTA astrocyte function increases stress-induced depression susceptibility
Summary
This study is the first to clearly define the protective role of VTA astrocytes in stress-related emotional disorders and to reveal a complete regulatory pathway: “Stress → Ca²⁺ elevation in VTA astrocytes → adenosine release → activation of A1 receptors on GABAergic interneurons → disinhibition of DA neurons → alleviation of emotional disorders.”
This finding breaks through the traditional neuron-centered research framework and provides astrocytes and the adenosine A1 receptor as two novel therapeutic targets with low addiction risk for stress-related psychiatric disorders such as anxiety and depression. It also lays an important theoretical foundation for developing safer and more precise treatment strategies.
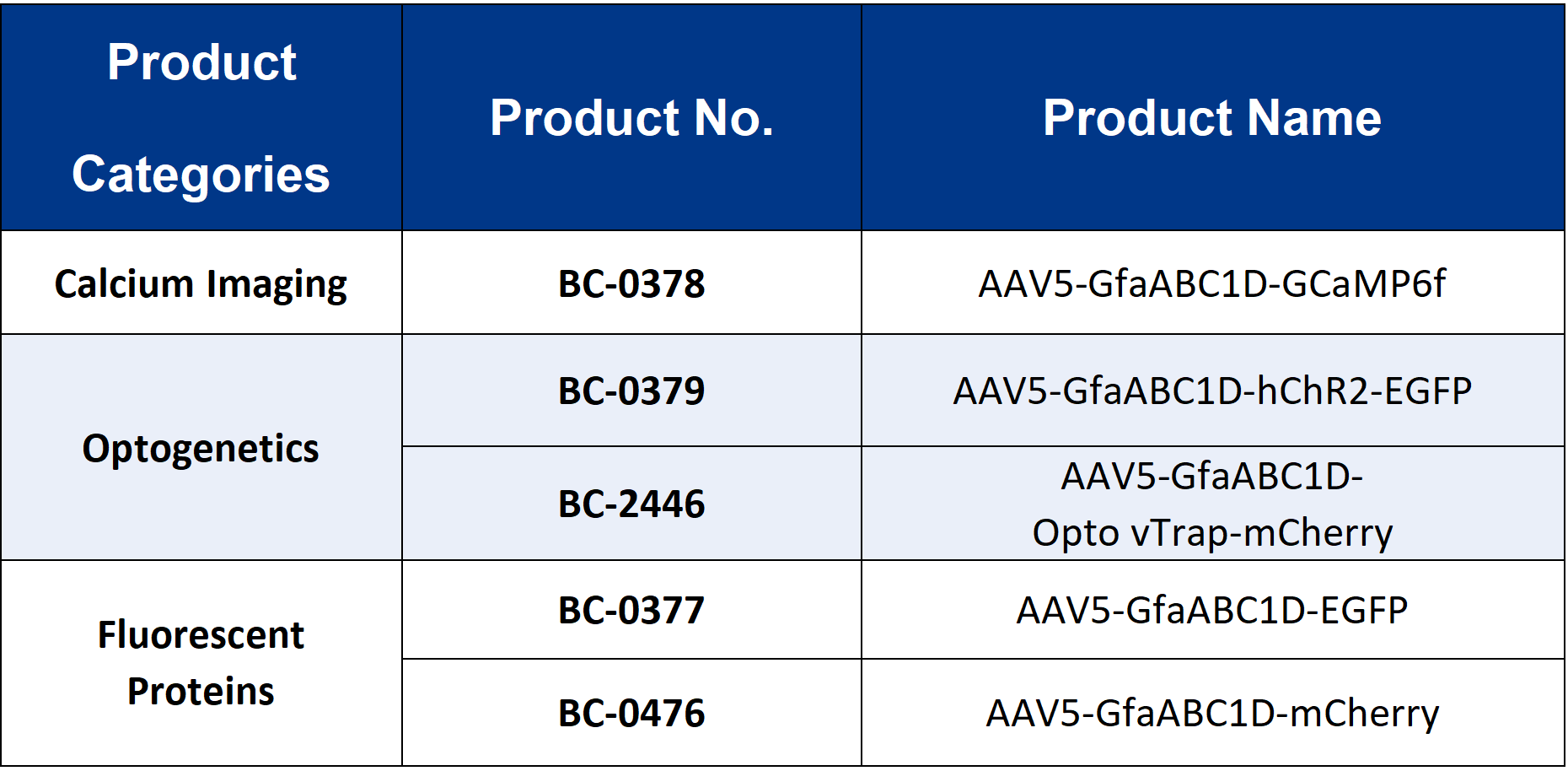
The viral tools used in this study were provided by Brain Case Biotech.
Researchers who are interested in this type of product are welcome
to contact us at bd@ebraincase.com