Client Article | Nat. Commun. | Lixin Xie & Qingjun Zhou's Team Reveals the Role and Mechanism of the Sympathetic Nervous System in Dry Eye Disease
Release time:2025-09-19 15:02:08
Tear secretion by the lacrimal gland and other structures is essential for maintaining ocular surface hydration and protection. Insufficient tear production leads to aqueous-deficient dry eye, with a global adult prevalence of 5%–50%. Traditionally, tear secretion has been considered primarily regulated by the parasympathetic nervous system (PSNS), while the role of the sympathetic nervous system (SNS) has remained controversial. Its in vivo pathological function and downstream pathways under dry eye stress conditions have not been clearly defined, hindering the development of effective treatments.
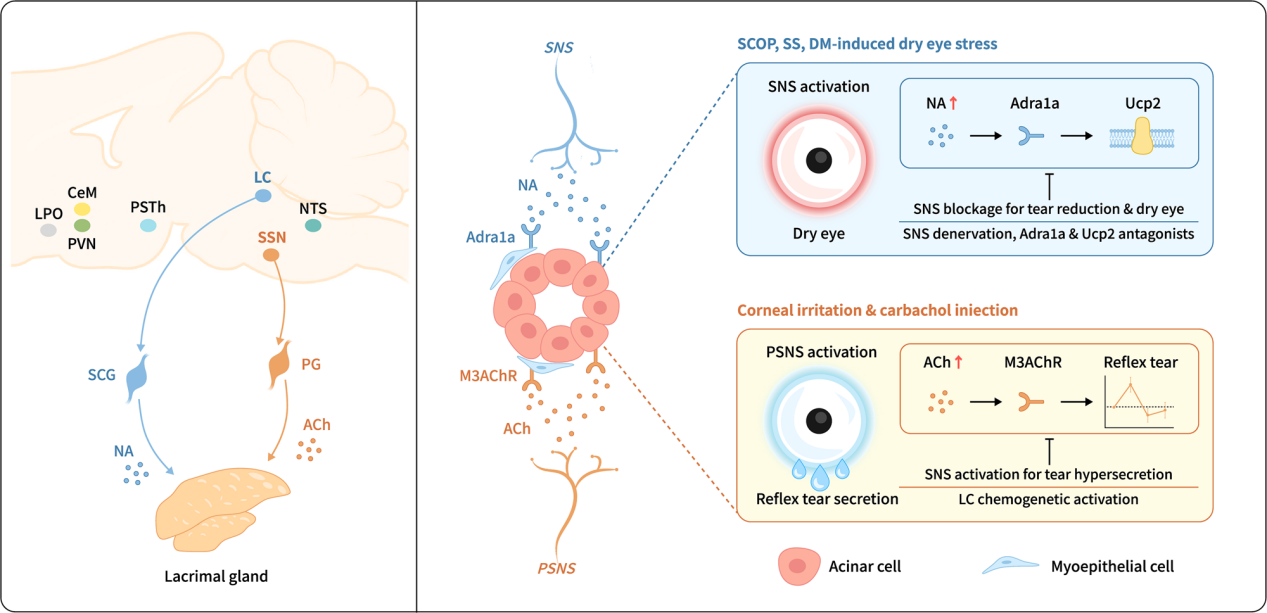
On June 5, 2025, a research team led by Academician Lixin Xie and Researcher Qingjun Zhou from the Ophthalmology Institute of Shandong First Medical University published a study in Nature Communications titled "A gatekeeper sympathetic control of lacrimal tear secretion and dry eye onset through the NA-Adra1a-Ucp2 pathway."The study revealed that the SNS innervates the lacrimal gland and is activated under various dry eye stress conditions. This activation leads to the release of norepinephrine (NA), which stimulates the α1a-adrenergic receptor (Adra1a) and regulates mitochondrial uncoupling protein 2 (Ucp2), ultimately reducing tear secretion and promoting the onset of dry eye.
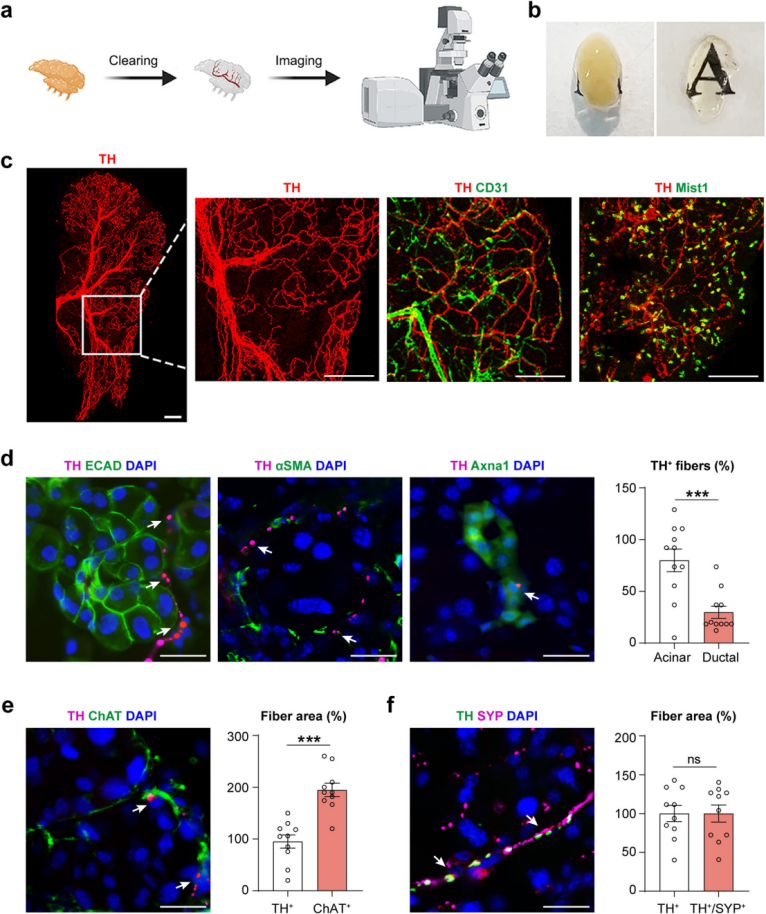
Sympathetic Innervation of the Mouse Lacrimal Gland
To visualize the complete neural network, the extraorbital lacrimal glands of mice were subjected to tissue clearing and fluorescent imaging (Figure 1a). Sympathetic nerve innervation was identified using immunolabeling of tyrosine hydroxylase (TH). After tissue clearing, the lacrimal glands became optically transparent (Figure 1b). This enabled visualization of the entire sympathetic nerve network and branching patterns at single-fiber resolution (Figure 1c).
Co-immunolabeling was then performed using TH along with the vascular endothelial marker CD31, acinar cell marker Mist1, epithelial marker E-cadherin (ECAD), myoepithelial cell marker αSMA, ductal cell marker Axna1, and parasympathetic nerve marker choline acetyltransferase (ChAT). Anatomically, sympathetic nerves in the lacrimal gland were classified into large bundles along blood vessels and fine fibers extending into acini and the surrounding stroma. Most fibers surrounded acinar and myoepithelial cells, with some located near ductal cells (Figure 1c–d). Quantitative analysis showed that the density of TH⁺ sympathetic nerves was about half that of ChAT⁺ parasympathetic nerves (Figure 1e), consistent with previous reports describing parasympathetic dominance in lacrimal gland innervation.
Furthermore, sympathetic nerves expressed synaptophysin (SYP), a marker of synaptic vesicles and presynaptic terminals (Figure 1f). These findings establish the distinct distribution patterns of sympathetic and parasympathetic innervation in the mouse lacrimal gland.
Figure 1. Visualization of sympathetic nerves in the mouse lacrimal gland
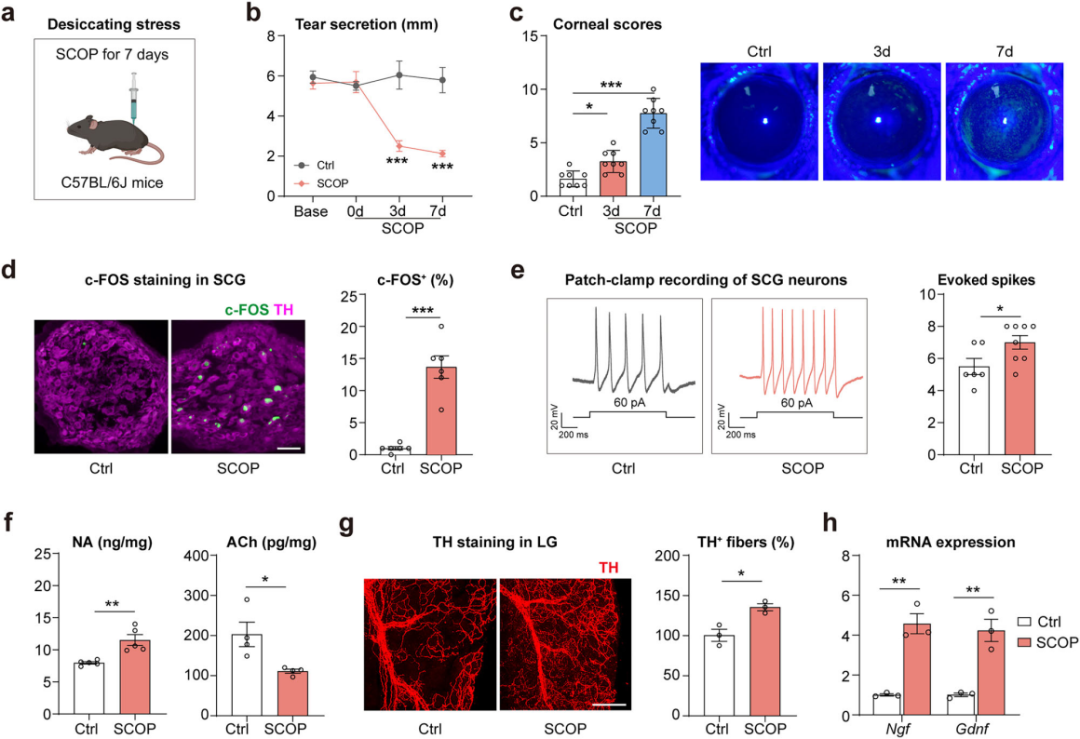
Dry Eye Stress Induces Sympathetic Nervous System Activation
The SNS plays a critical role in the “fight-or-flight” response. To investigate changes in SNS activity under dry eye stress, a mouse model of dry eye was established using scopolamine (SCOP) combined with chronic desiccating stress (Figure 2a). Results showed that after 3 days of SCOP treatment, tear secretion decreased (Figure 2b), and corneal damage appeared after 7 days (Figure 2c).
At the same time, the number of c-FOS⁺ TH⁺ neurons in the superior cervical ganglion (SCG)—the largest sympathetic ganglion—in dry eye mice increased 12-fold compared to controls (Figure 2d). Patch-clamp recordings demonstrated elevated action potential frequency in SCG neurons (Figure 2e).
Furthermore, after 7 days of SCOP treatment, NA levels and sympathetic nerve density in the lacrimal gland were increased (Figure 2f–g), while acetylcholine (ACh) levels and ChAT expression were decreased (Figure 2f). Additionally, the expression of neurotrophic factors NGF and GDNF was upregulated (Figure 2h).
Together, these findings indicate that SCOP and desiccating stress can activate sympathetic innervation in the lacrimal gland, suggesting its involvement in reduced tear secretion and the development of dry eye.
Figure 2. Dry eye stress induces sympathetic nervous system activation
Sympathetic Nervous System Activation Suppresses Tear Secretion Norepinephrine Reduces Tear Secretion in Dry Eye Mice
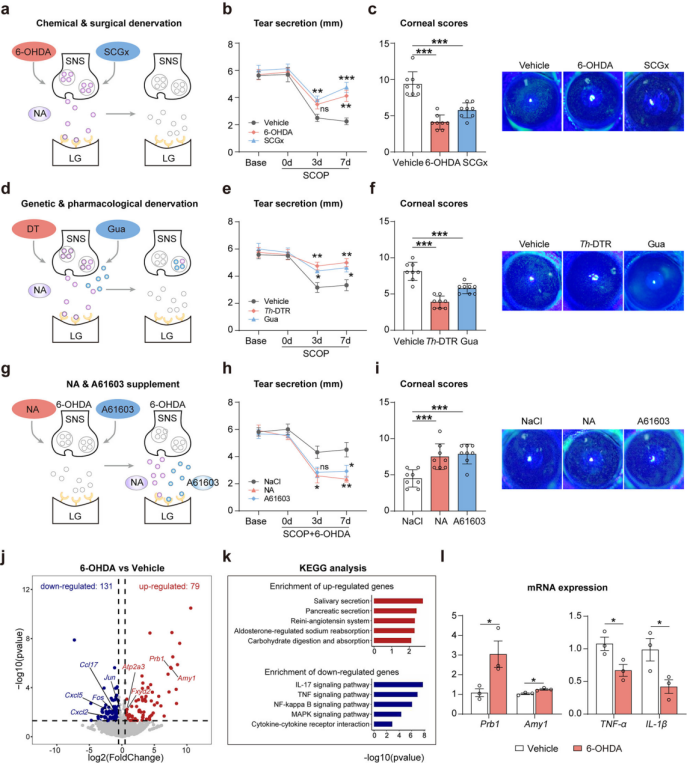
To clarify the role of SNS activation in regulating lacrimal tear secretion, sympathetic denervation was induced in SCOP-treated dry eye mice using chemical (6-OHDA), surgical (superior cervical ganglionectomy, SCGx), and genetic (Th-DTR mice) approaches (Figure 3a). These treatments led to reduced SNS innervation density and NA levels in the lacrimal gland. Both 6-OHDA and SCGx significantly increased tear secretion and improved corneal damage (Figure 3b–c).
For targeted ablation of sympathetic nerves, Th-Cre mice were crossed with ROSA26i-DTR mice to generate Th-DTR mice, in which diphtheria toxin (DT) selectively eliminates TH⁺ neurons and fibers (Figure 3d). DT treatment reduced SNS density and NA concentration in the lacrimal gland. After exposure to SCOP and desiccating stress, Th-DTR mice showed significantly improved tear secretion (Figure 3e) and reduced corneal damage (Figure 3f). Similar results were observed in mice pretreated with guanethidine, a compound that blocks NA release from sympathetic nerve terminals (Figure 3d–f). Overall, sympathetic denervation or blockade promotes tear secretion in dry eye mice.
To verify the direct role of SNS-derived NA in tear secretion regulation, NA or the Adra1a agonist A61603 was administered to 6-OHDA-pretreated dry eye mice (Figure 3g). Compared to vehicle-treated controls, NA infusion or A61603 injection re-induced the dry eye phenotype, with reduced tear secretion (Figure 3h) and aggravated corneal damage (Figure 3i), resembling untreated dry eye mice. Adrenalectomy ruled out contributions from adrenal-derived NA, confirming that SNS-derived NA directly causes tear reduction in dry eye mice.
To directly link enhanced tear secretion from sympathetic denervation with restored lacrimal gland function, researchers performed transcriptomic and histological analyses. In 6-OHDA–treated glands, 79 genes were upregulated and 131 were downregulated (Figure 3j). KEGG analysis showed enrichment of upregulated genes in secretion-related pathways and downregulated genes in inflammatory pathways (Figure 3k). qPCR confirmed increased transcription of Prb1 (encoding proline-rich salivary protein 1) and Amy1 (encoding amylase), and decreased expression of TNF-α and IL-1β (Figure 3l).In summary, inactivating the SNS enhances tear secretion by improving lacrimal gland function in dry eye mice.
Figure 3. Inactivation of the SNS increases tear secretion in dry eye mice
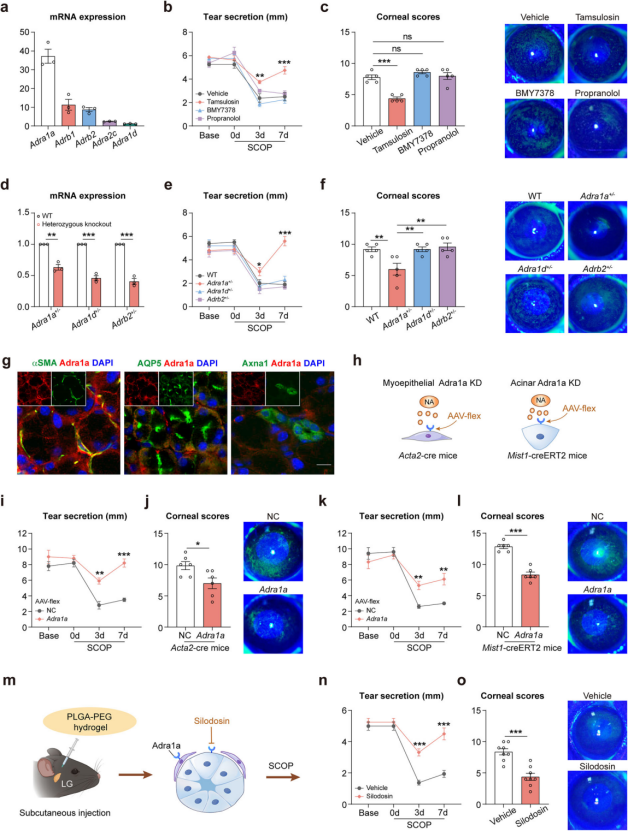
Adra1a, Not Adra1d, Mediates Sympathetic Nervous System–Induced Tear Reduction
The sympathetic nervous system regulates tear secretion via NA acting on adrenergic receptors. mRNA analysis of mouse lacrimal glands revealed that Adra1a was the most abundantly expressed subtype (Figure 4a). To determine the functional receptor involved, dry eye mice were pretreated with various antagonists: tamsulosin (Adra1a antagonist), BMY7378 (Adra1d antagonist), and propranolol (β1- and β2-AR antagonist). Only tamsulosin significantly improved tear secretion and reduced corneal damage (Figure 4b–c).
Further confirmation came from heterozygous knockout mouse models (Figure 4d): only Adra1a⁺/⁻ mice—not Adra1d⁺/⁻ or Adrb2⁺/⁻ mice—showed enhanced tear secretion (Figure 4e) and alleviated corneal damage (Figure 4f). Together, both pharmacological and genetic evidence demonstrate that Adra1a, rather than Adra1d or Adrb2, plays a key role in SNS-mediated tear reduction.
Building on these findings, the researchers explored the downstream mechanism of sympathetic regulation. Immunolabeling revealed that Adra1a colocalizes with both myoepithelial and acinar cells in the lacrimal gland (Figure 4g). Conditional knockdown of Adra1a using Acta2-cre and Mist1-creERT2 mice (Figure 4h) significantly improved dry eye symptoms (Figure 4i–l). qPCR analysis of purified cell types showed that tamsulosin upregulated genes involved in tear production in acinar cells and tear secretion in myoepithelial cells. These results indicate that the sympathetic nervous system regulates both tear generation and secretion by targeting acinar and myoepithelial cells.
Finally, local hydrogel-based delivery of the Adra1a antagonist silodosin (Figure 4m) significantly improved tear secretion and reduced corneal damage in dry eye mice (Figure 4n–o).
In conclusion, upon activation, the sympathetic nervous system releases NA, which binds to Adra1a receptors on acinar and myoepithelial cells, thereby suppressing tear production and secretion under dry eye stress.
Figure 4. Adra1a mediates the dominant effect of the SNS on tear secretion
Adra1a Blockade Is Effective Across Multiple Models of Aqueous-Deficient Dry Eye
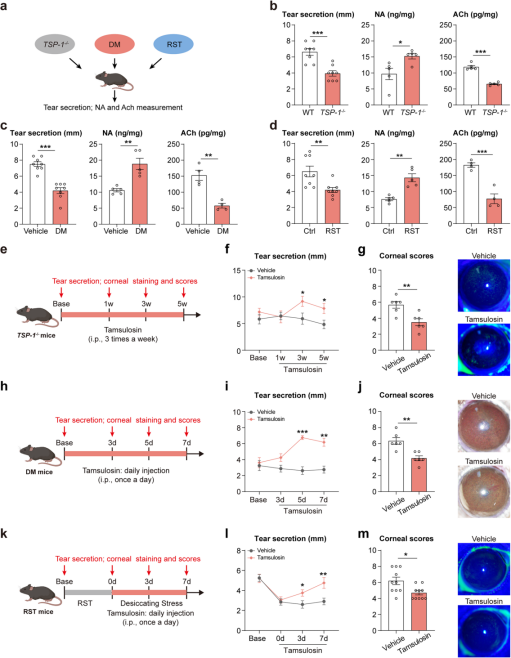
Dry eye disease is a multifactorial condition, and lacrimal gland dysfunction can lead to aqueous-deficient dry eye. To validate the role of SNS activation in different types of aqueous-deficient dry eye, researchers examined three distinct mouse models: Sjögren’s syndrome (TSP-1⁻/⁻), diabetes, and restraint stress. All three models exhibited significant reductions in tear secretion and corneal damage (Figure 5a–d). Compared to control and wild-type mice, these dry eye models also showed a similar pattern of increased NA and decreased ACh levels in the lacrimal gland (Figure 5a–d).
Intraperitoneal injection of the Adra1a antagonist tamsulosin (Figure 5e, h, k) significantly improved tear secretion (Figure 5f, i, l) and reduced corneal damage (Figure 5g, j, m) across all three models. Notably, improvement in tear secretion in TSP-1⁻/⁻ mice was slower compared to the diabetic and restraint stress models.
In summary, these results suggest that activation of the sympathetic nervous system in the lacrimal gland may represent a common pathological mechanism and therapeutic target across multiple forms of aqueous-deficient dry eye.
Figure 5. Adra1a blockade is effective across multiple models of aqueous-deficient dry eye in mice
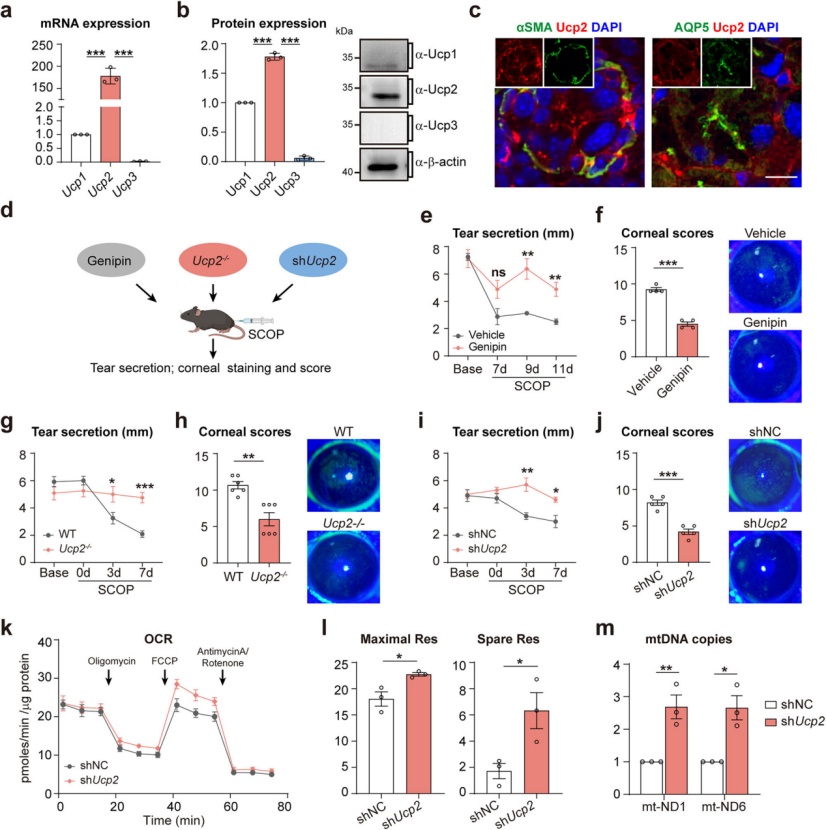
After confirming the dominant role of Adra1a, the study investigated the downstream pathways by which SNS activation regulates tear secretion. Uncoupling protein 2 (Ucp2), known to be activated by the SNS and involved in various physiological processes, was found to be highly expressed in the mouse lacrimal gland (Figure 6a–b). Both scopolamine and chronic desiccating stress significantly upregulated Ucp2 expression. Immunolabeling revealed that Ucp2 colocalizes with myoepithelial and acinar cells (Figure 6c).
To determine the role of Ucp2 in SNS-mediated tear regulation, dry eye was induced in C57BL/6J mice pretreated with the Ucp2 inhibitor genipin or using Ucp2⁻/⁻ knockout mice (Figure 6d). In both models, tear secretion (Figure 6e–g) and corneal damage (Figure 6f–h) were significantly improved.
Furthermore, injection of an AAV encoding shRNA targeting Ucp2 (shUcp2) into the lacrimal gland also enhanced tear secretion and reduced corneal damage in dry eye mice (Figure 6i–j). This knockdown increased mitochondrial respiratory capacity and mitochondrial DNA copy number (Figure 6k–m). In summary, Ucp2 in the lacrimal gland plays a key role in mediating SNS-regulated tear secretion under chronic dry eye stress.
The Locus Coeruleus Participates in Sympathetic Nervous System–Mediated Tear Reduction
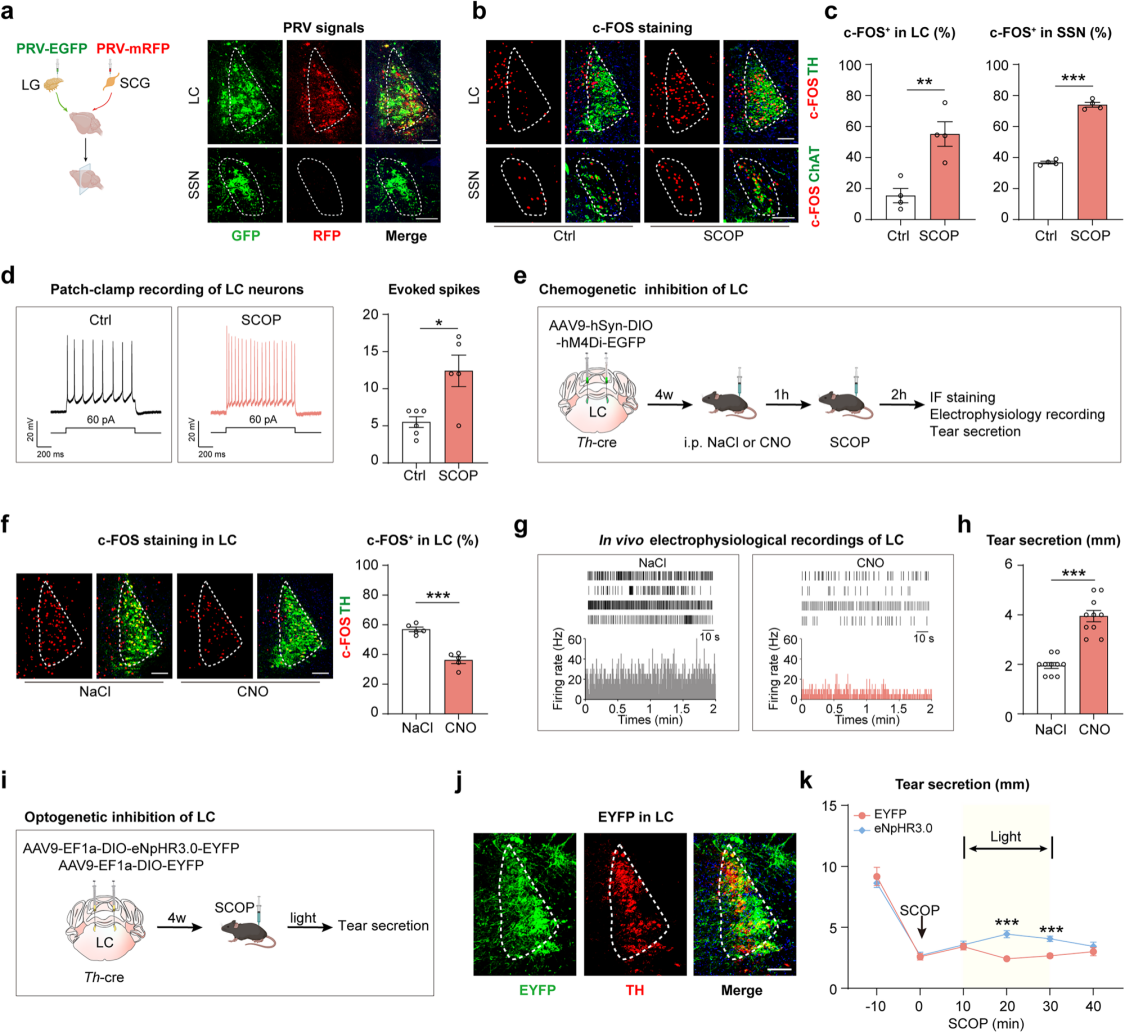
To identify the brain regions that send norepinephrine-releasing (noradrenergic) outputs to the lacrimal gland, researchers conducted retrograde trans-synaptic tracing by injecting PRV viruses expressing EGFP and mRFP into the lacrimal gland and the SCG, respectively (Figure 7a). On day 2 after PRV injection, GFP signals were detected in the superior salivatory nucleus (SSN), a known source of parasympathetic innervation to the lacrimal gland. On day 3, dual-labeled GFP and mRFP signals were clearly observed in the locus coeruleus (LC), nucleus of the solitary tract (NTS), and paraventricular nucleus (PVN); by day 4, signal intensity increased and was detected in additional brain regions.
Most dual-labeled neurons in the LC expressed TH, while neurons in the SSN—exclusively labeled with GFP—expressed ChAT, indicating that the SSN sends parasympathetic projections, while the LC, NTS, and PVN provide sympathetic input to the lacrimal gland. The LC has previously been reported to regulate systemic sympathetic output. To explore its role in tear secretion, mice treated with scopolamine showed increased numbers of c-FOS⁺ cells in both the LC and SSN (Figure 7b–c), and LC neurons exhibited increased burst firing frequency (Figure 7d), indicating activation of both sympathetic LC and parasympathetic SSN neurons.
To test the functional role of the LC, researchers used chemogenetics (injecting AAV9 encoding hM4Di into the LC of Th-Cre mice) (Figure 7e–g) and optogenetics (injecting AAV9 encoding eNpHR3.0) (Figure 7i–j) to inhibit noradrenergic neurons in the LC. Inhibition of the LC resulted in a significant increase in tear secretion in Th-Cre mice (Figure 7h, k). In summary, the LC is a key sympathetic brain nucleus involved in regulating tear secretion from the lacrimal gland.
Figure 7. The locus coeruleus participates in SNS-mediated tear reduction
Locus Coeruleus–Superior Cervical Ganglion–Lacrimal Gland Axis Regulates Reflex Tear Secretion
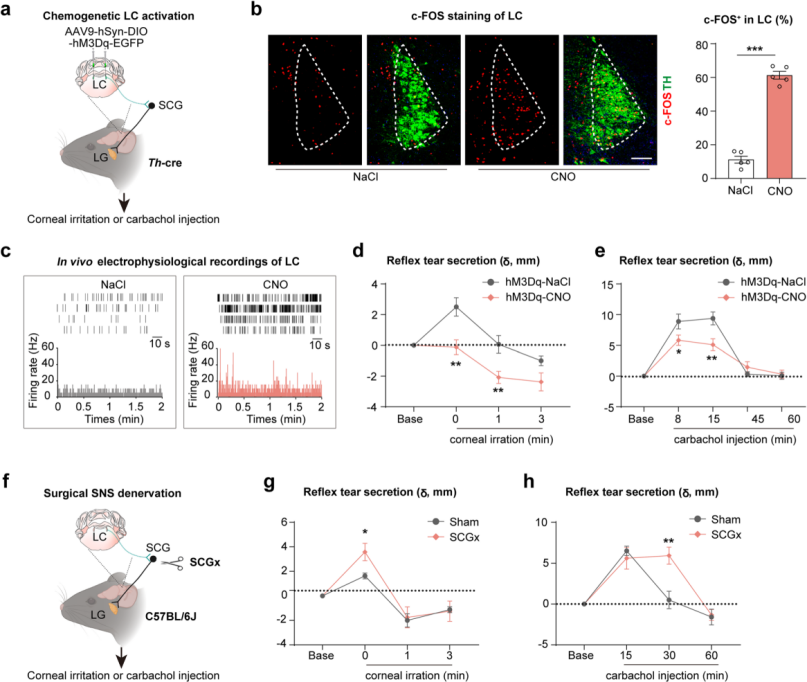
To further investigate the role of the locus coeruleus–superior cervical ganglion–lacrimal gland (LC–SCG–LG) axis in the regulation of reflex tear secretion, mouse models were established using corneal nylon thread stimulation and intraperitoneal injection of carbachol, both of which induce rapid reflex tearing. AAV9-DIO-hM3Dq was bilaterally injected into the LC of Th-cre mice (Fig. 8a). Immunofluorescence staining and electrophysiological recordings confirmed LC neuronal activation after CNO administration (Fig. 8b–c). Results showed that reflex tear secretion induced by corneal stimulation was completely suppressed in this model (Fig. 8d), while secretion induced by carbachol injection was partially reduced (Fig. 8e).
Furthermore, after peripheral sympathetic nerve ablation via SCGx(Fig. 8f), corneal stimulation elicited even greater reflex tear secretion (Fig. 8g), and carbachol-induced tear secretion exhibited a prolonged duration (Fig. 8h). In summary, the LC–SCG–LG axis precisely regulates both the magnitude and duration of reflex tear secretion, serving to prevent excessive tearing.
Figure 8 | Reflex Tear Secretion Induced by Activation and Inhibition of the Sympathetic Nervous System (SNS)
Conclusion
This study visualized the complete sympathetic and parasympathetic innervation network within the mouse lacrimal gland and, for the first time, revealed the regulatory role of the sympathetic nervous system as a physiological "gatekeeper" in vivo. It identified the locus coeruleus–superior cervical ganglion–lacrimal gland axis (LC–SCG–LG axis) as a key pathway controlling tear secretion in response to both dry eye stress and reflex stimulation. These findings not only deepen our understanding of the mechanisms underlying tear secretion regulation but also offer a novel theoretical framework and potential therapeutic targets for the prevention and treatment of dry eye disease.
All viral vectors used in this study are available from Brain Case Biotech(bd@ebraincase.com):