慢性社交应激会导致社交功能受损,但其影响社交行为的具体神经机制尚不明确。慢性情绪应激(CES)可使小鼠出现社交回避、社交新奇偏好丧失等表型,而岛叶皮层与前额叶皮层均参与社交与情绪调控,且前额叶富含催产素受体,可选择性调节社交记忆,但其在应激中的作用仍不清楚。

2026年3月26日,军事医学科学院吴海涛教授团队在《Neuron》期刊上发表题为Dual anterior insula-prefrontal cortex circuits mediate chronic stress-induced social interaction deficits的研究论文。该研究揭示慢性应激通过双环路损害社交行为:前岛叶背侧皮层(AID)内谷氨酸能阳性神经元构成的AIDCaMKIIα→前额叶联合皮层(FrA)通路过度激活可驱动社交恐惧,并通过兴奋前岛叶小清蛋白(PV)阳性中间神经元,对前岛叶腹侧皮层(AIV)投射至边缘前皮层(PrL)的环路产生侧向抑制,同时下调PrL区催产素受体信号,共同导致社交新奇偏好缺陷。抑制AID→FrA环路、激活AIV→PrL环路、给予催产素受体激动剂或抑制PV中间神经元均可逆转社交缺陷。该研究不仅阐明了慢性应激破坏社交行为的双环路整合机制,也为社交焦虑、抑郁等精神障碍相关社交退缩提供了精准的环路与分子治疗靶点。
本研究采用替代性应激模型对雄性C57BL/6J小鼠进行连续10天CES处理,使其每日观察入侵鼠被CD‑1小鼠攻击10–15分钟,该范式使观察鼠仅承受间接情绪应激,无任何身体接触;对照组小鼠饲养条件相同但无入侵鼠(图1A)。依据社交互动测试(SIT)中的社交互动比率将应激小鼠分为易感型(SUS)与耐受型(RES),SUS小鼠表现出明显社交回避,对CD‑1小鼠嗅探时间显著减少,RES小鼠社交偏好与对照组无显著差异(图1B-D);社交新奇偏好测试(SNPT)显示各组小鼠均具备正常社交能力,而仅SUS小鼠对陌生小鼠无社交新奇偏好,提示其存在社交新奇辨别能力缺陷(图1E‑G);对40个脑区c‑Fos表达图谱的检测发现,对照组较笼养基线小鼠有5个社交情绪相关脑区激活升高,SUS与RES小鼠的前额叶皮层、基底外侧杏仁核、终纹床核、腹内侧下丘脑等应激焦虑相关脑区均进一步激活,且二者在前岛叶皮层(aIC)呈现差异模式,即SUS小鼠AID激活增强、AIV激活减弱,RES小鼠AID与AIV均激活增强(图1H、I)。
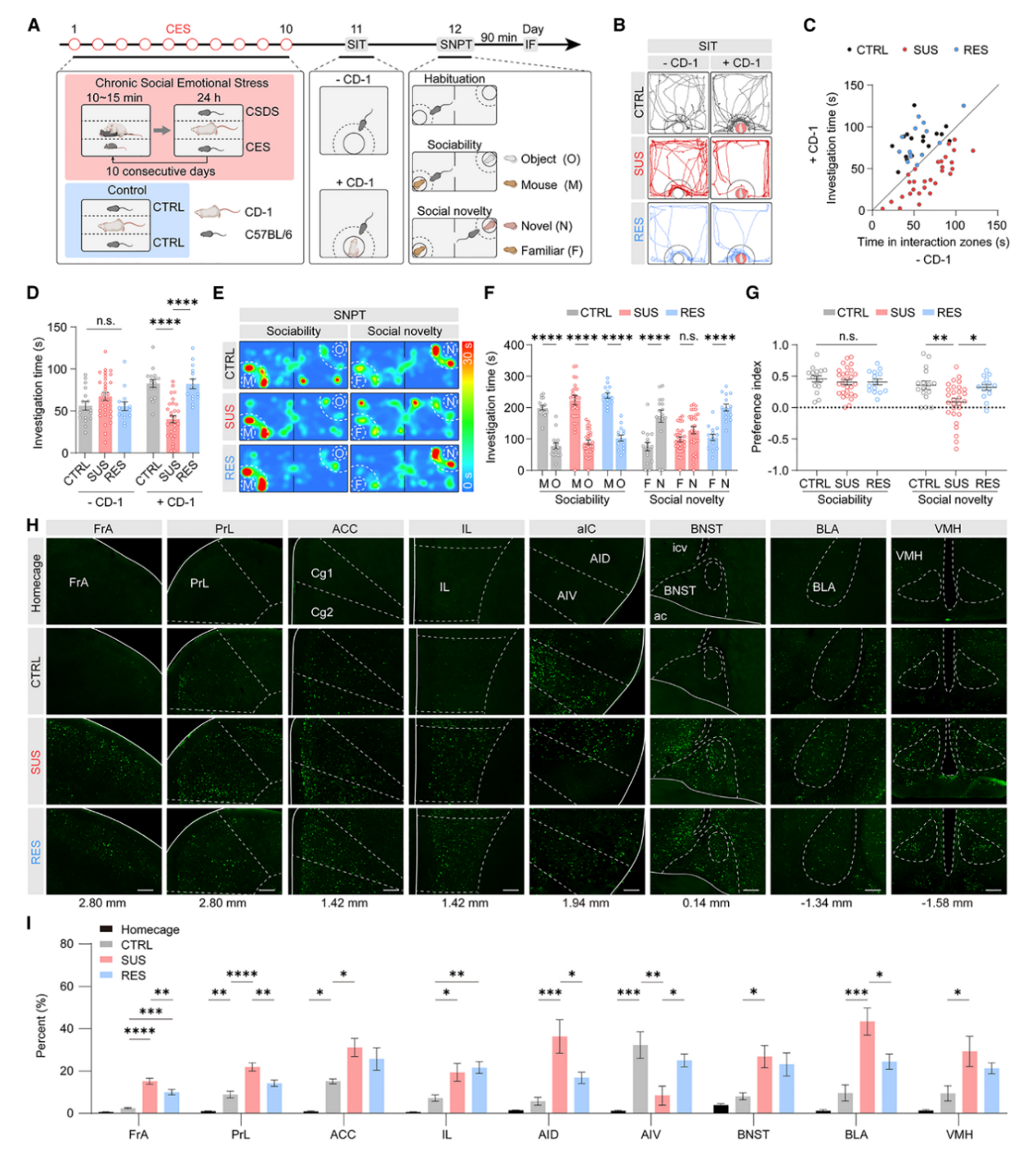
图1.CES诱导SUS小鼠出现社交恐惧与社交新奇性损伤
研究通过fMOST技术重构AID与AIV脑区26个神经元的树突和轴突形态(图2A),这些神经元包含CaMKIIα谷氨酸能兴奋性神经元与GABA能抑制性中间神经元,全脑投射广泛;对比艾伦脑图谱发现,AID与AIV神经元投射模式整体符合前岛叶皮层连接特征,但在前额叶存在明显差异,AID神经元主要投射至前额叶联合皮层(FrA),AIV神经元则主要投射至边缘前皮层(PrL)(图2B)。通过向PrL与FrA注射逆行示踪剂验证(图2C),结果显示投射至FrA的神经元多分布于AID,投射至PrL的神经元多分布于AIV,且均集中于皮层2/3层,深层标记极少(图2D-F),两者交叉投射比例不足5%,为两条平行独立的神经通路。结合CaMKIIα-Cre::Ai9、Viaat-Cre::Ai9报告基因小鼠与CTB-488逆行示踪实验,证实这两类投射神经元主要为CaMKIIα阳性的兴奋性神经元。最终鉴定出AIDCaMKIIα→FrA与AIVCaMKIIα→PrL两条分离通路,提示AID和AIV可通过各自特异的投射模式调控社交行为。
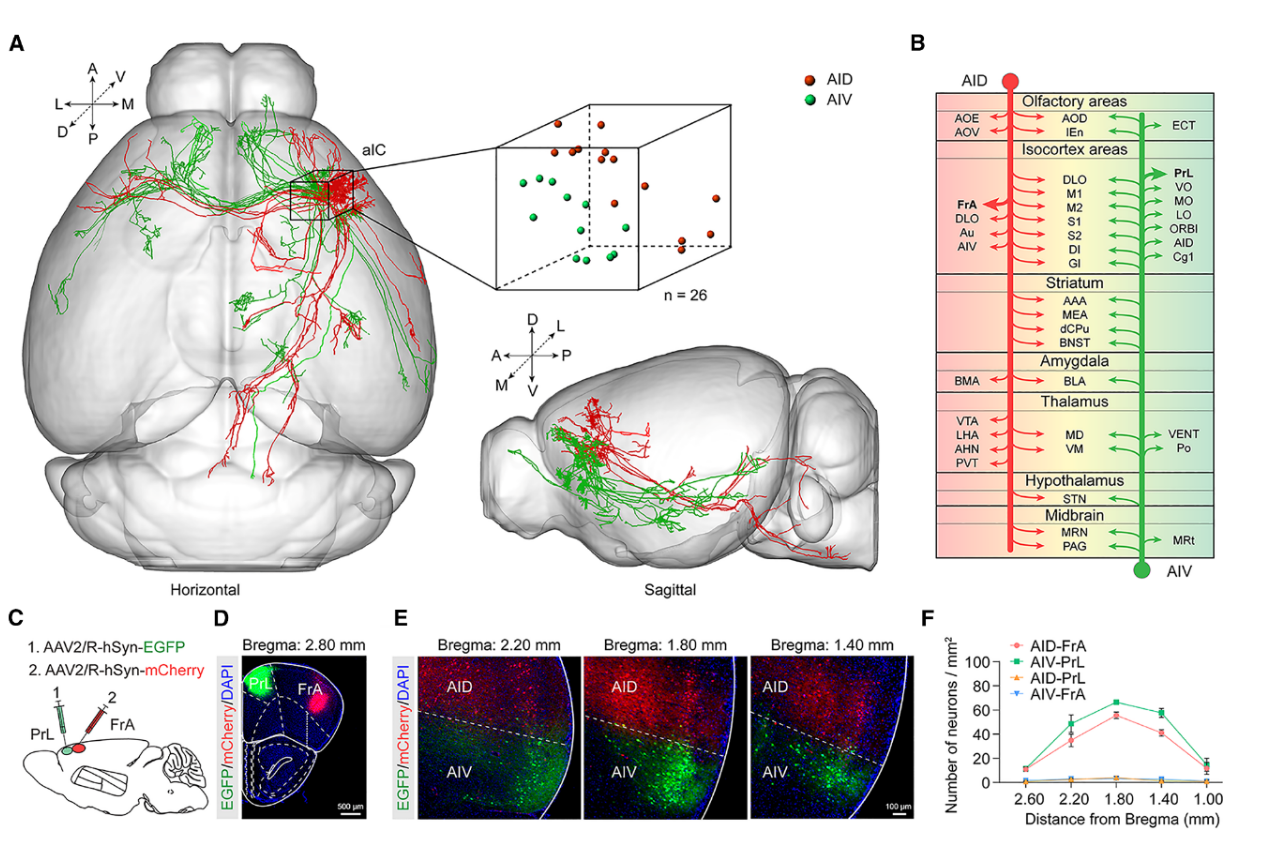
图2.AID与AIV神经元的亚型及投射模式
CES可使SUS小鼠出现社交恐惧与社交新奇偏好受损,为阐明其动态机制,研究采用光纤光度法在体记录AIDCaMKIIα→FrA与AIVCaMKIIα→PrL两条神经环路在社交应激及互动过程中的活动变化(图3A、B)。将逆行钙指示剂病毒分别注射至FrA或PrL脑区,同时在AID与AIV上方植入光纤(图3C-F)。两条环路表现出显著的功能分离:SUS小鼠的AIDCaMKIIα→FrA神经元仅在靠近恐惧化CD-1小鼠时异常激活(图3G-K),介导社交回避;而AIVCaMKIIα→PrL通路在面对陌生小鼠时活动降低(图3L-P),导致社交新奇偏好受损。综上,aIC→PFC投射网络通过环路特异性重构调控社交行为。
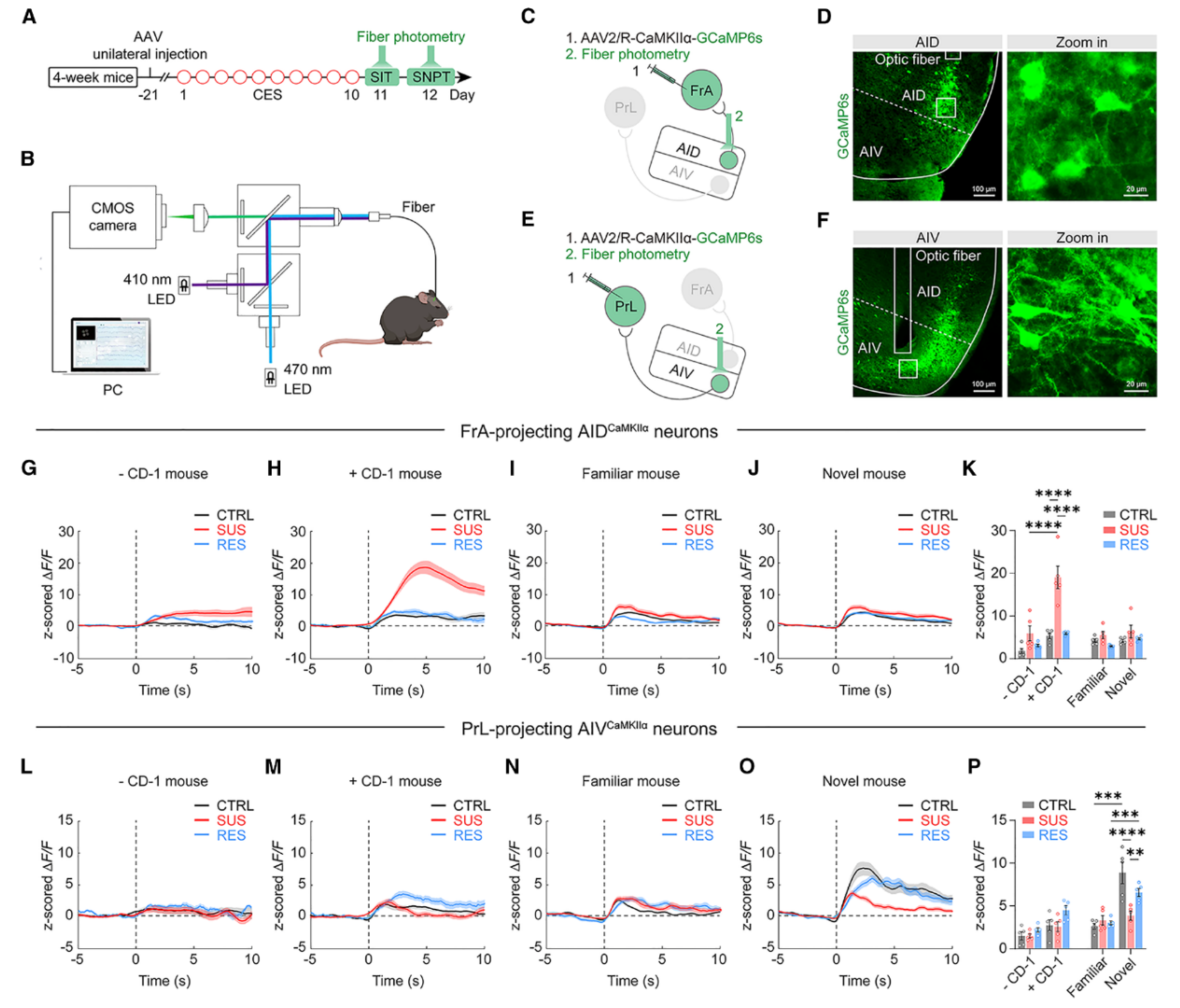
图3. 社交情境下AIDCaMKIIα→FrA与AIVCaMKIIα→PrL神经元的活动变化
进一步采用化学遗传学与光遗传学对AIDCaMKIIα→FrA与AIVCaMKIIα→PrL进行双向操控(图4A、B,4G、H)。结果显示,通过光遗传抑制AIDCaMKIIα→FrA环路可同时改善社交回避与新奇偏好(图4C-F);而10Hz激活AIVCaMKIIα→PrL环路则特异性恢复社交新奇偏好,不影响恐惧相关行为(图4I-N)。作者发现通过化学遗传抑制正常小鼠AIVCaMKIIα→PrL环路可选择性地损伤社交新奇偏好,但不影响新物体识别与运动能力。综上,CES诱发的社交恐惧依赖AID→FrA环路过度激活,社交新奇偏好缺陷则由AIV→PrL环路活动不足导致,证实两条aIC→PFC环路的功能分离与失衡是应激相关社交障碍的核心机制。
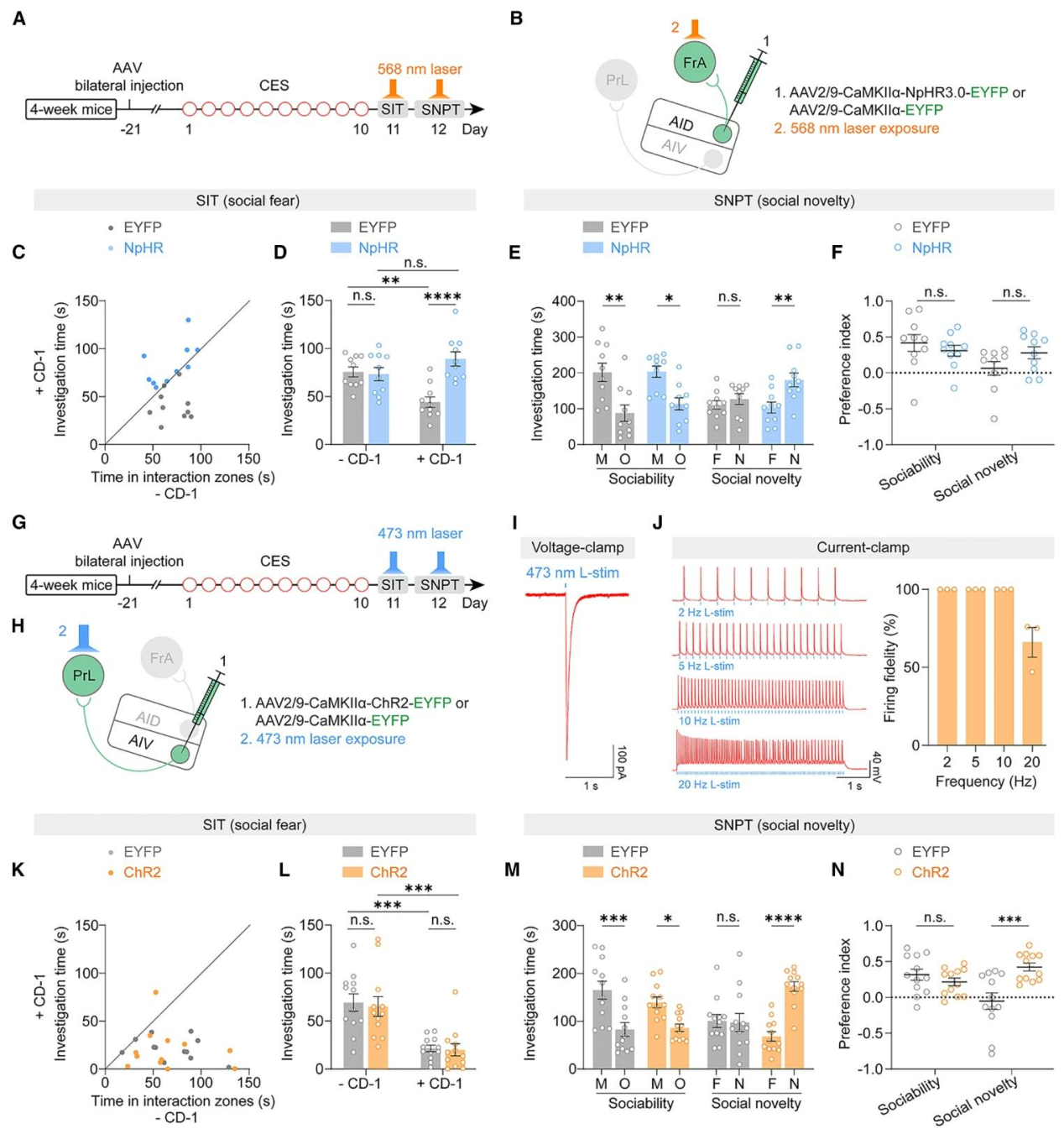
图4.光遗传学操控AIDCaMKIIα→FrA与AIVCaMKIIα→PrL环路可双向调控应激诱导的社交功能障碍
研究通过顺行示踪发现,AIV主要支配PrL的生长抑素阳性(SST)GABA中间神经元,且CES后SUS小鼠PrLGABA能神经元中催产素受体Oxtr表达选择性降低。为验证PrLOXTR神经元(受AIV支配且表达OXTR)对社交新奇偏好的编码作用,在AIV注射AAV2/1-Cre、在PrL注射Cre依赖且Oxtr启动子驱动的GCaMP6s病毒,特异性记录PrLOXTR神经元活动(图5A、B),结果发现对照组小鼠探究陌生同类时Ca²⁺反应显著,SUS小鼠则明显减弱(图5E-G),免疫染色证实约90%的GCaMP6s阳性神经元共表达OXTR(图5C、D)。利用光遗传学激活该通路可恢复SUS小鼠社交新奇偏好,不影响基础社交能力(图5H-M)。病毒介导CRISPR-Cas9在投射至PrL的AIV神经元中特异性敲低OXTR,未产生明显神经毒性(图5N、O),OXTR免疫反应性降低约70%(图5P、Q),并选择性导致社交新奇偏好缺陷而不改变基础社交互动(图5R、S),证实OXTR不可或缺。综上,OXTR表达下调损害PrL中受AIV输入的SST中间神经元功能,是社交新奇偏好缺陷的核心机制,也明确了AIV→PrLOXTR环路的关键调控作用。
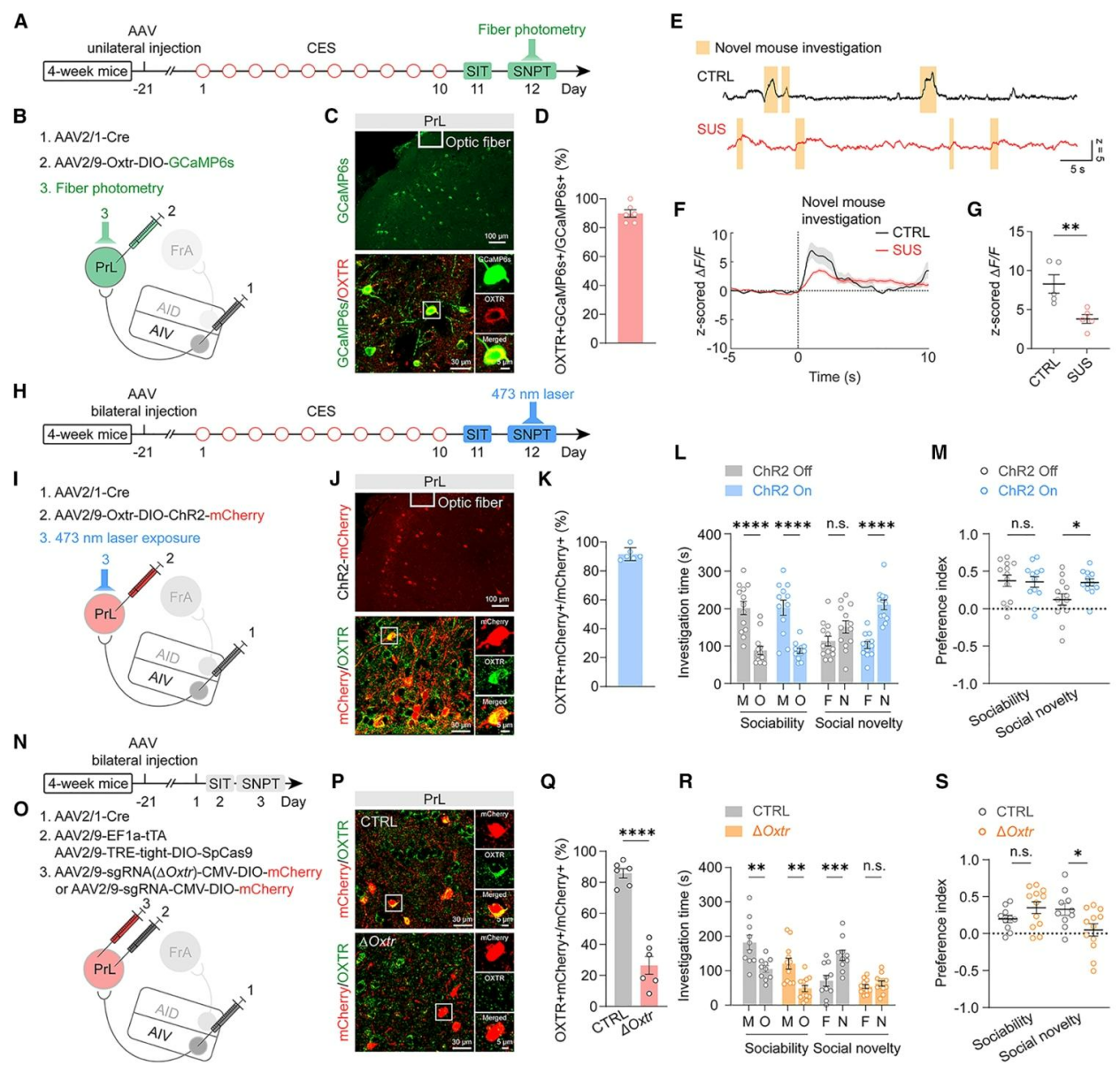
图5. AIVCaMKIIα→PrL环路中的OXTR信号调控社交新奇偏好
为明确PrLOXTR神经元的催产素能输入来源及其对AIV→PrL通路的调控,在Oxtr-Cre小鼠采用逆行跨单突触示踪(图6A),发现PrLOXTR神经元的直接突触前输入主要来自下丘脑室旁核(PVN),视上核(SON)仅有少量标记,且PVN中大部分标记神经元为催产素阳性(图6B-D)。在Oxtr-Cre小鼠AIV注射AAV-ChR2-EGFP,PrL注射Cre依赖的AAV-DIO-mCherry,分别标记AIV末梢和PrLOXTR神经元(图6E、F),急性脑片记录显示,灌流催产素受体激动剂TGOT可增强AIV→PrL突触的光诱发兴奋性突触后电流(oEPSC)幅值,拮抗剂阿托西班则起抑制作用(图6G、H);同时TGOT可提高配对脉冲比率,阿托西班则降低该比率(图6I、J),提示OXTR激活可提升突触前释放概率、调控突触短时程可塑性。在体实验中,向SUS小鼠双侧PrL微量注射TGOT(图6K、L),可显著增加其对陌生小鼠的探究时间(图6M、N),证实直接激活PrL区OXTR即可恢复SUS小鼠的社交新奇偏好。
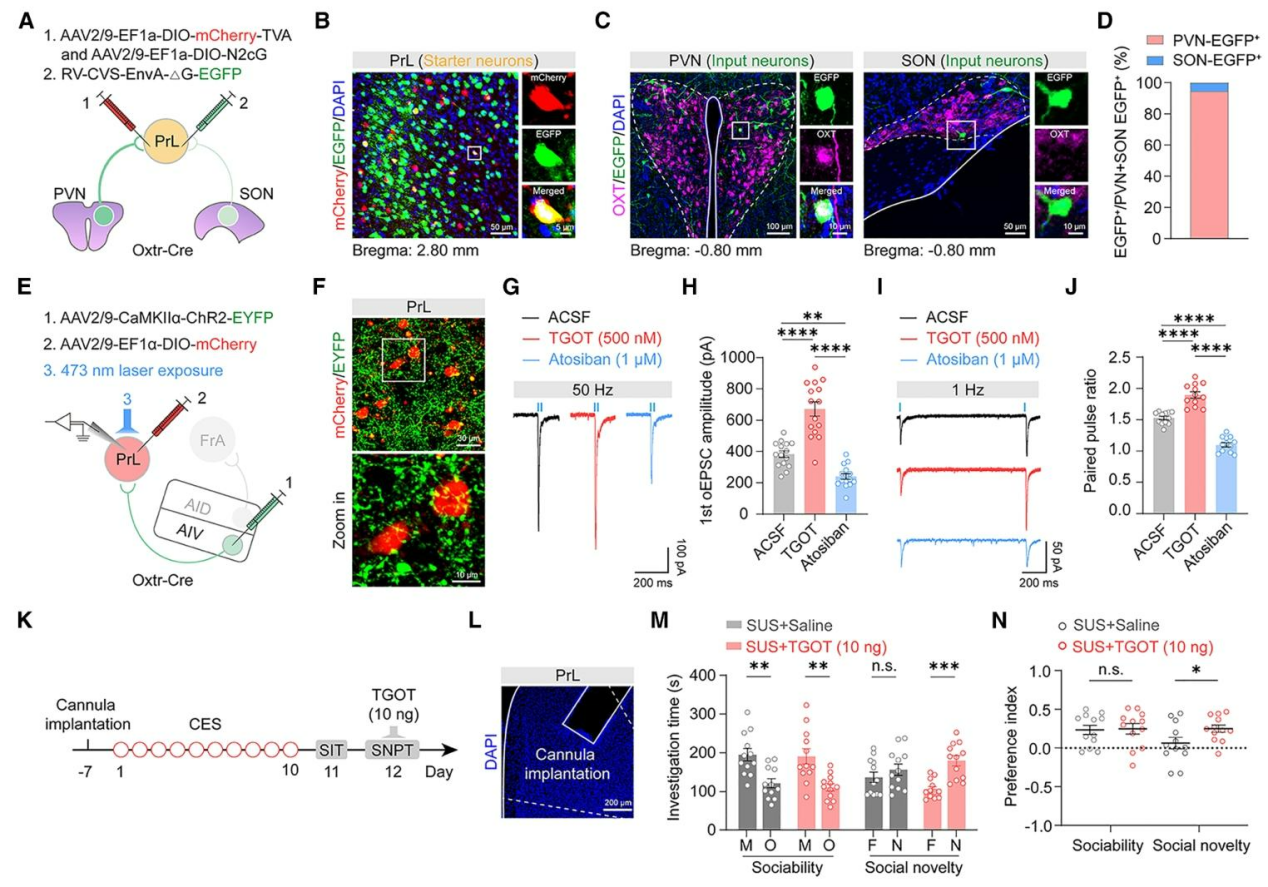
图6. 来自PVN的催产素能输入调控AIVCaMKIIα→PrLOXTR环路的突触效能,进而调节社交行为
基于岛叶局部抑制机制,研究推测SUS小鼠中过度激活的AID→FrA环路通过aIC局部GABA神经元抑制AIV→PrL通路,并综合运用多种技术进行验证。
利用RV逆行示踪技术,在Viaat-Cre小鼠双侧PrL注射逆行 RV-helper,三周后于AIV区注射跨突触狂犬病毒(RV),在aIC区标记出起始投射神经元(TagBFP+mCherry+),并通过Cre依赖的EGFP信号特异性标记其上游GABA能抑制性输入,实现该抑制性环路的精准可视化(图7A、B)。FrA区注射逆行AAV2/R-CaMKIIα-ChR2,特异性标记AIDCaMKIIα→FrA神经元;光刺激aIC区可在aICGABA中间神经元上诱发出兴奋性突触后电位,提示存在功能性输入(图7C),光诱发电流可被1μM TTX阻断、20mM 4-AP恢复,证实为单突触连接(图7D-F)。在支配AIVCaMKIIα→PrL神经元的aICGABA能中间神经元中,83%可接收AIDCaMKIIα→FrA神经元输入,17%无反应(图7D),表明AIDCaMKIIα→FrA神经元活动可驱动aICGABA能中间神经元从而抑制AIVCaMKIIα→PrL神经元。
SUS小鼠中,支配AIV→PrL的aICGABA中间神经元内在兴奋性显著升高(图7G、H),其中93%(14/15)表现为快放电表型(FS)(图7I、J),提示其主为PV+ 抑制性中间神经元。为验证上述结论,在PV-Cre小鼠中利用跨单突触狂犬病病毒,在支配AIVCaMKIIα→PrL神经元的aICPV中间神经元上特异性表达ChR2-EGFP(图7K),约90%的EGFP阳性神经元与PV共定位(图7L、M),光激活该类细胞可在AIVCaMKIIα→PrL投射神经元诱发抑制性突触后电流(IPSC),且可被GABA受体拮抗剂印防己毒素完全阻断,证实其抑制作用(图7N、O);光纤光度记录显示,SUS小鼠在新奇社交探索时,该aICPV中间神经元的Ca²⁺反应显著增强(图7P-T),提示其抑制驱动增强导致AIVCaMKIIα→PrL环路活动低下;而通过光抑制(eNpHR3.0)相同神经元群时可显著恢复小鼠的社交新奇偏好(图7W、X)。综上,aICPV中间神经元的活动介导了对AIVCaMKIIα→PrL环路的侧向抑制,导致SUS小鼠出现社交新奇偏好缺陷。
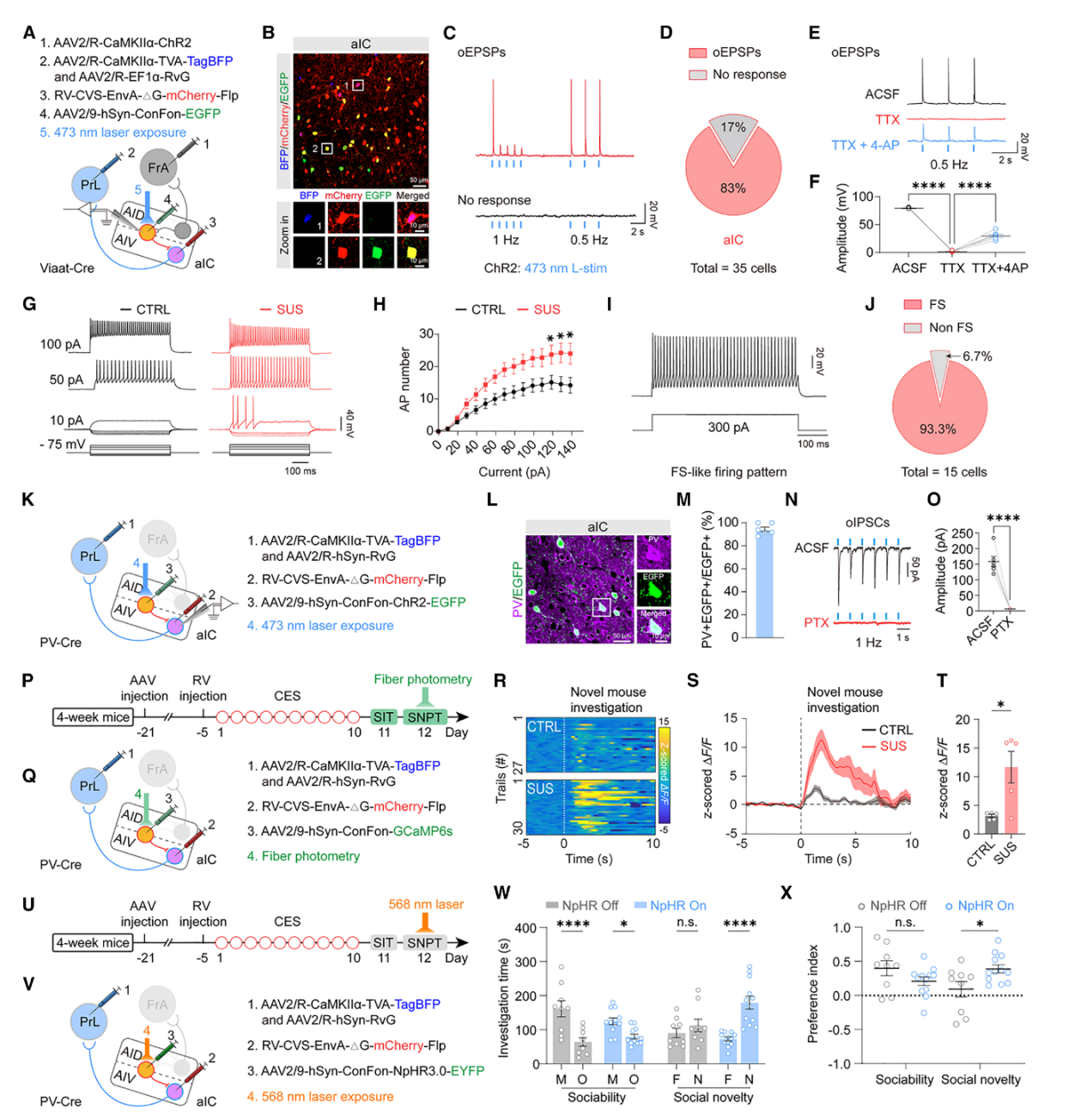
图7. SUS小鼠中过度激活的aICPV微环路介导AIDCaMKIIα→FrA环路对 AIVCaMKIIα→PrL环路的抑制作用
研究通过CES小鼠模型,发现aIC内存在两条功能相反的并行神经环路:AID→FrA环路介导社交恐惧与回避,AIV→PrL环路调控社交新奇偏好;慢性应激会使AID→FrA过度激活,并通过aICPV中间神经元对AIV→PrL产生侧向抑制,同时导致PrL脑区OXTR信号下调,共同造成社交回避与社交新奇偏好缺失;而通过光遗传、化学遗传或药物激活AIV→PrL环路、增强OXTR信号均可有效逆转应激导致的社交缺陷。该研究从环路、细胞和分子层面完整阐释了慢性压力导致社交障碍的神经机制,为相关精神疾病的治疗提供了精准靶点。
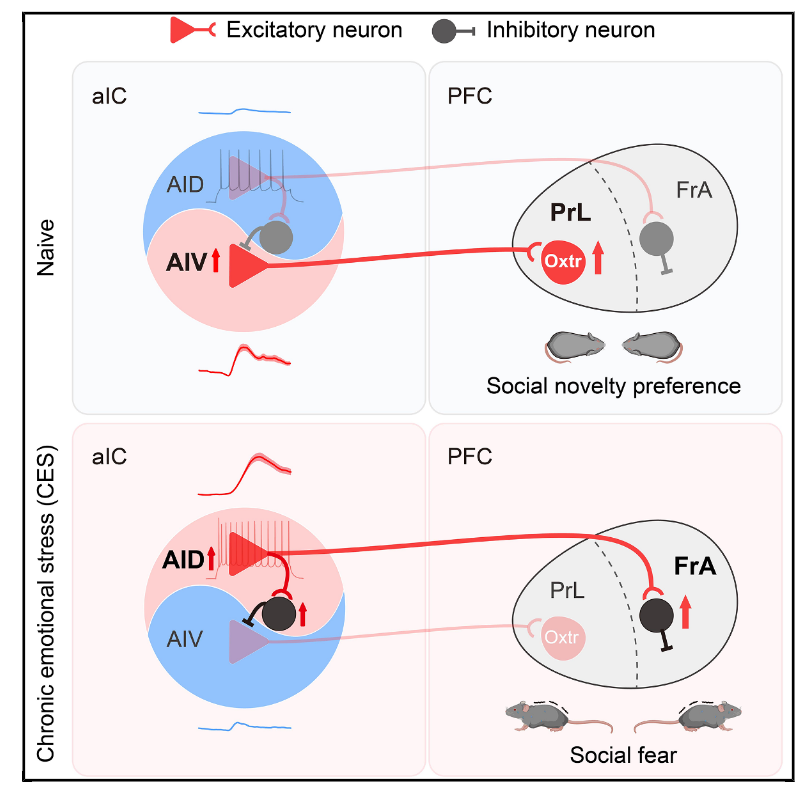
图8. 总结图
本文使用的工具病毒布林凯斯均可提供:
同时布林凯斯也可提供各类定制服务请联系小布:18971216876(微信同号)或者咨询所在区域的销售经理获取更多信息。





