迷走神经含80%的传入纤维,是内脏活动的重要调节神经,但在内脏痛感知中的定性功能尚未明确;TRPV1⁺神经元介导疼痛和炎症反应,DRG中该神经元参与胰腺炎痛,但NG中其作用未知。孤束核(NTS)是内脏传入信号的核心中继站,下丘脑室旁核(PVN)参与自主神经和痛觉情绪调控,二者的神经连接及在急性胰腺炎(AP)急性内脏痛中的作用尚未验证。
2026年3月22日,中山大学医学院徐婷团队在《Brain, Behavior, and Immunity》发表题为“The vagus nerve activated by inflammation co-transmits acute visceral pain via the NTS-PVN pathway”相关文章,聚焦急性胰腺炎诱导的内脏痛神经传导机制,通过多学科技术明确了迷走神经介导的胰腺-大脑痛觉通路,为内脏痛的治疗提供了全新的神经环路靶点。

作者通过腹腔注射雨蛙素构建小鼠急性胰腺炎(AP)模型(图1A),结果显示,AP组血清淀粉酶显著高于生理盐水组且随给药天数上升(图1B);AP组胰腺小叶间纤维间隔增厚水肿,伴淋巴细胞、中性粒细胞浸润(图1C),髓过氧化物酶(MPO)活性随给药次数升高(图1D)。检测发现,AP组胰腺组织三种关键炎症因子IL-1β、IL-6、TNF-α的mRNA及蛋白表达均显著上调(图1E、1F)。行为学检测中,AP组扭体阳性反应增多(图1G),提示存在自发性内脏痛;0.02g和0.4g冯·弗雷纤维丝刺激下,AP组缩退反应差异显著(图1H、1I),故选取这两个参数用于后续实验。综上,AP诱导的内脏痛模型构建成功。因随给药天数增加小鼠状态变差,后续实验均采用造模3天的小鼠。
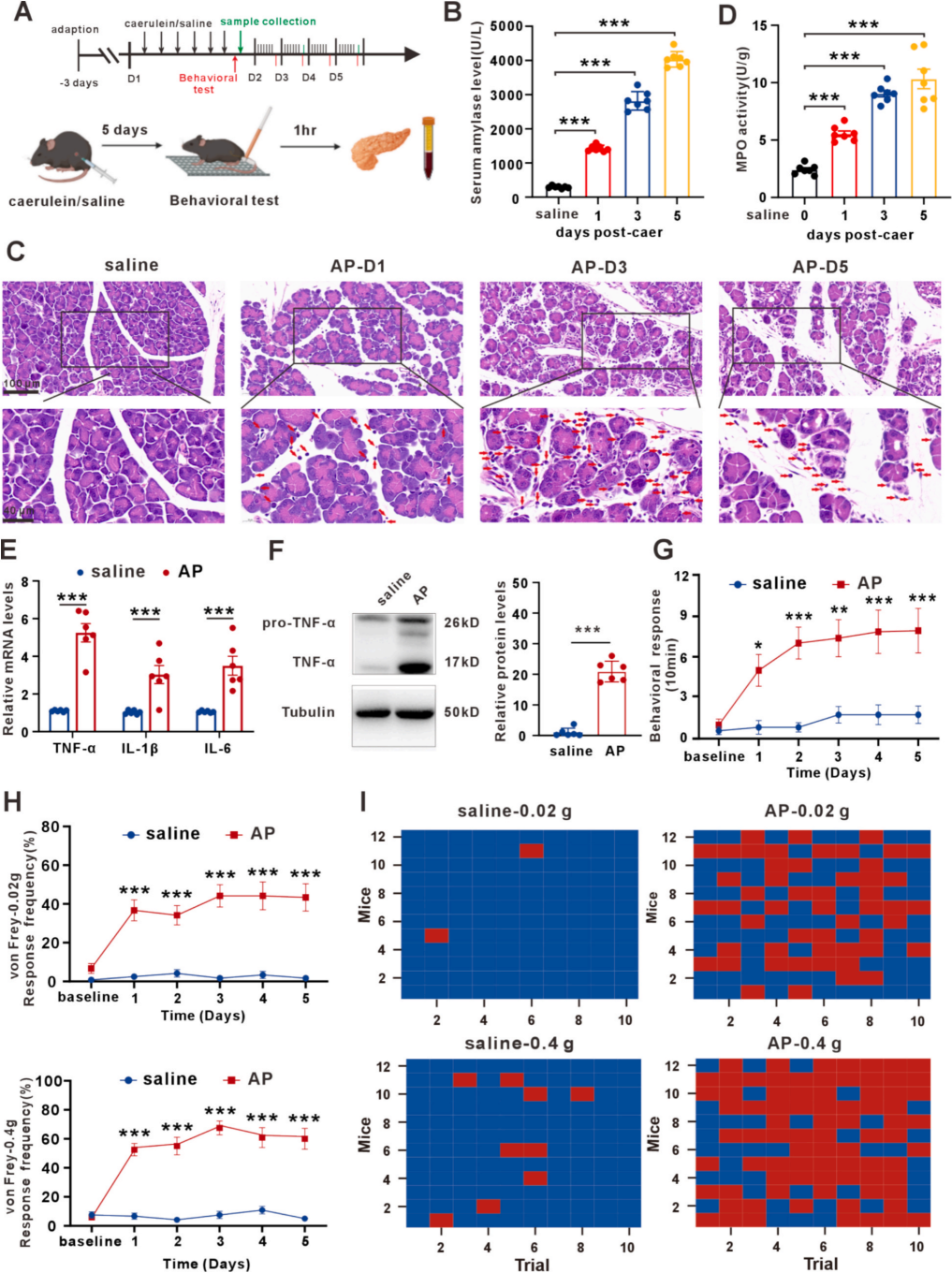
图1 雨蛙素诱导小鼠胰腺炎症与内脏痛表型
内脏感觉传导主要通过迷走神经与脊神经通路。用逆行示踪病毒注射胰腺标记胰腺支配神经元(图2A),结果显示结状神经节(NG)标记率为9.78±1.01%,背根神经节(DRG)标记率1.39%~8.91%,其中胸9段DRG最高(图2B-2D)。为验证示踪特异性,采用胰管灌注法,NG标记率12.10±1.83%,胸9段DRG最高。顺行示踪进一步证实,NG注射病毒后,胰腺腺泡及胰岛周边可见标记纤维。迷走神经是否介导内脏痛存在争议,此为研究核心。磷酸化细胞外信号调节激酶(pERK)是NG神经元激活标志物,cFos是中枢敏化指标。急性胰腺炎(AP)组小鼠NG的pERK、NTS的cFos信号均显著升高。双侧膈下迷走神经切断术(VGX)可显著降低二者信号(图2E、2F),并缓解AP小鼠痛觉行为(图2G-2H)。为了明确VGX对AP病理的影响,检测发现其可改善胰腺病理、血清淀粉酶、MPO活性及促炎因子表达。采用双病毒策略(图2I),特异性消融胰腺投射的NG神经元,可显著缓解AP小鼠内脏痛(图2J-2L)。综上,迷走神经在胰腺至大脑的内脏痛传导中起主导作用,NG→NTS轴为核心传导通路。
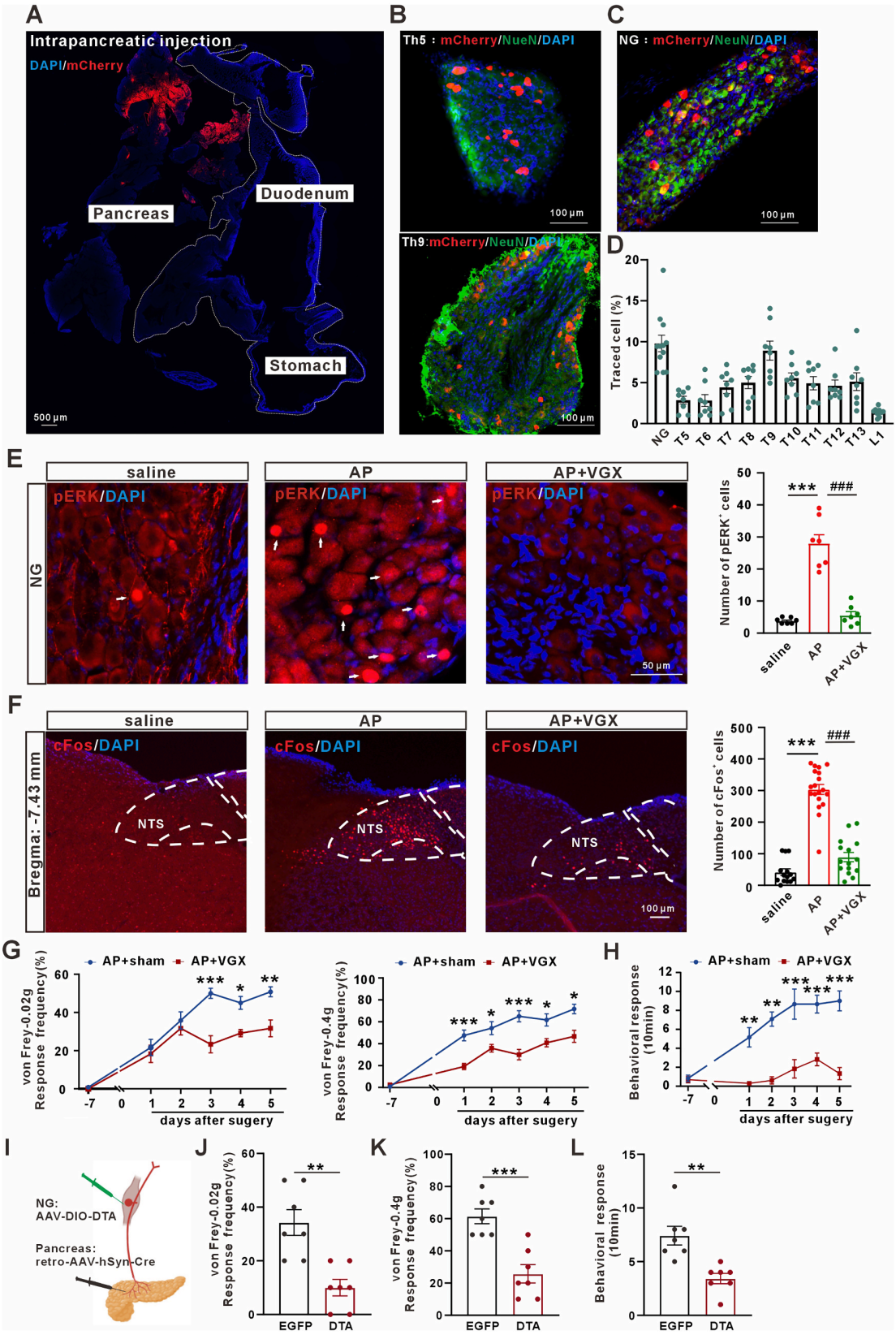
图2 迷走神经介导急性胰腺炎内脏痛传入
作者分析Kupari, J.等的数据集发现,NG细胞可聚类为17个亚群、两大集群,TRPV1特异性高表达于第二集群(NG12-NG18,占比更高)(图3A);该集群特异性基因的GO分析显示其与痛觉感知相关(图3B)。据此推测NG的TRPV1⁺神经元介导急性胰腺炎内脏痛传入。免疫荧光共染色证实,胰腺注射逆行示踪病毒标记的NG神经元多为TRPV1阳性(图3C),且与AP诱导的pERK共定位。采用树脂毒素(RTX)清除双侧NG的TRPV1⁺神经元后,AP小鼠的机械刺激反应和扭体反应显著缓解(图3D、3E)。原代NG神经元电生理实验显示,AP中升高最显著的肿瘤坏死因子α(TNF-α)可降低神经元基强度电流、增加动作电位发放(图3F、3G),而TNF-α抑制剂SPD304可消除该效应(图3H),表明TNF-α可敏化NG神经元。
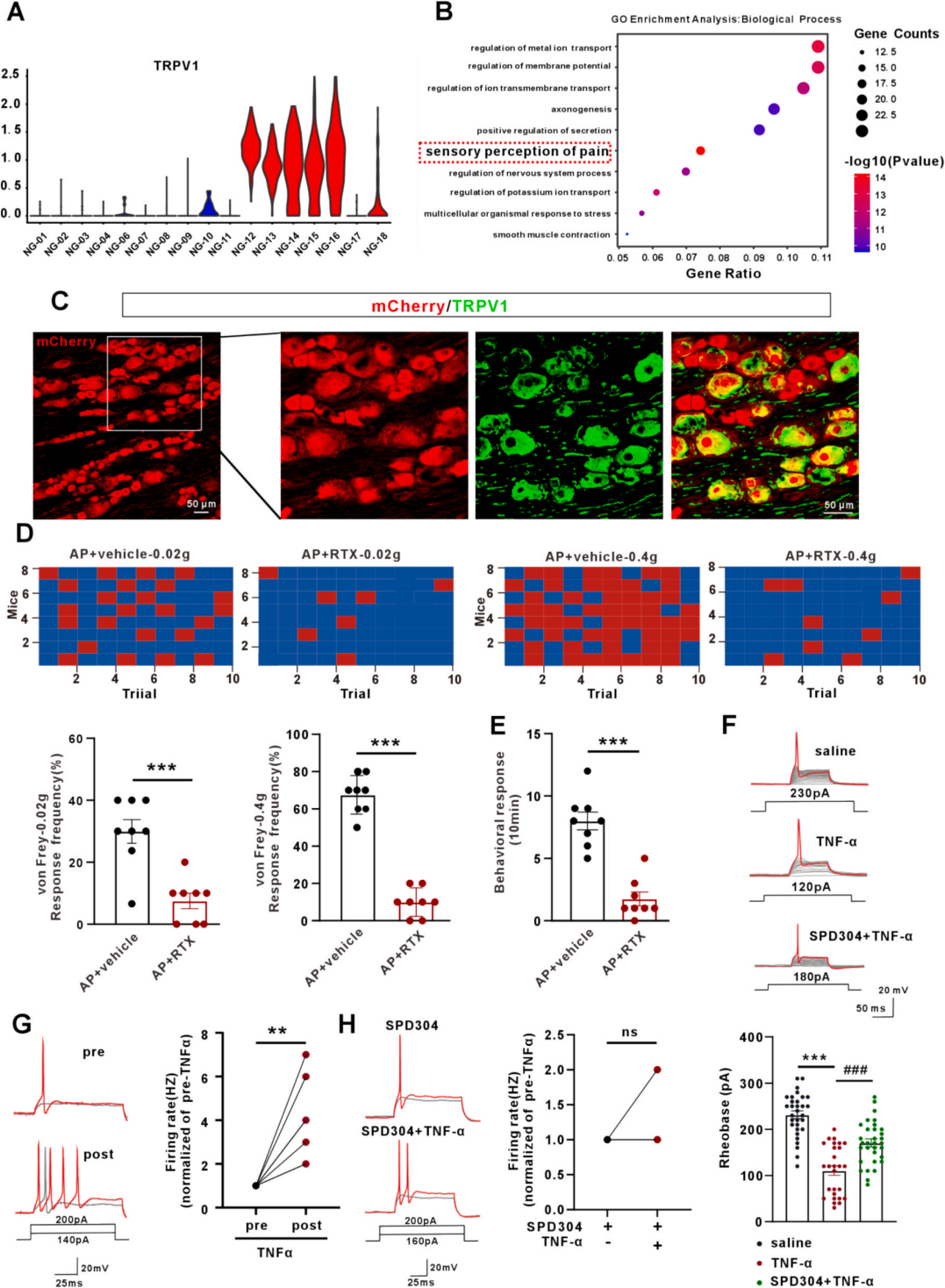
图3 结状神经节 TRPV1⁺神经元激活参与急性胰腺炎内脏痛
已有结果证实,炎症激活的迷走神经TRPV1神经元参与急性胰腺炎内脏痛传入,且NTS内cFos表达显著升高,切断迷走神经可逆转该痛觉反应与cFos激活。为明确NTS神经元激活是否由迷走神经TRPV1神经元介导,采用活性依赖示踪策略联合RV逆行示踪(图4A),鉴定出AP激活的NTS起始细胞(图4B),且这些神经元接收NG投射,NG内EGFP⁺信号为TRPV1神经元(图4C)。提示NTS激活神经元接收NG内TRPV1神经元传入;胰腺内检测到RV标记的EGFP⁺信号(图4D),证实提示胰腺与NTS之间存在从外周到中枢的神经环路。采用Gi‑DREADD化学遗传抑制胰腺→NG→NTS环路(图4E),结果显示抑制NG神经元活性可显著缓解AP小鼠内脏痛行为(图4F-4H)。
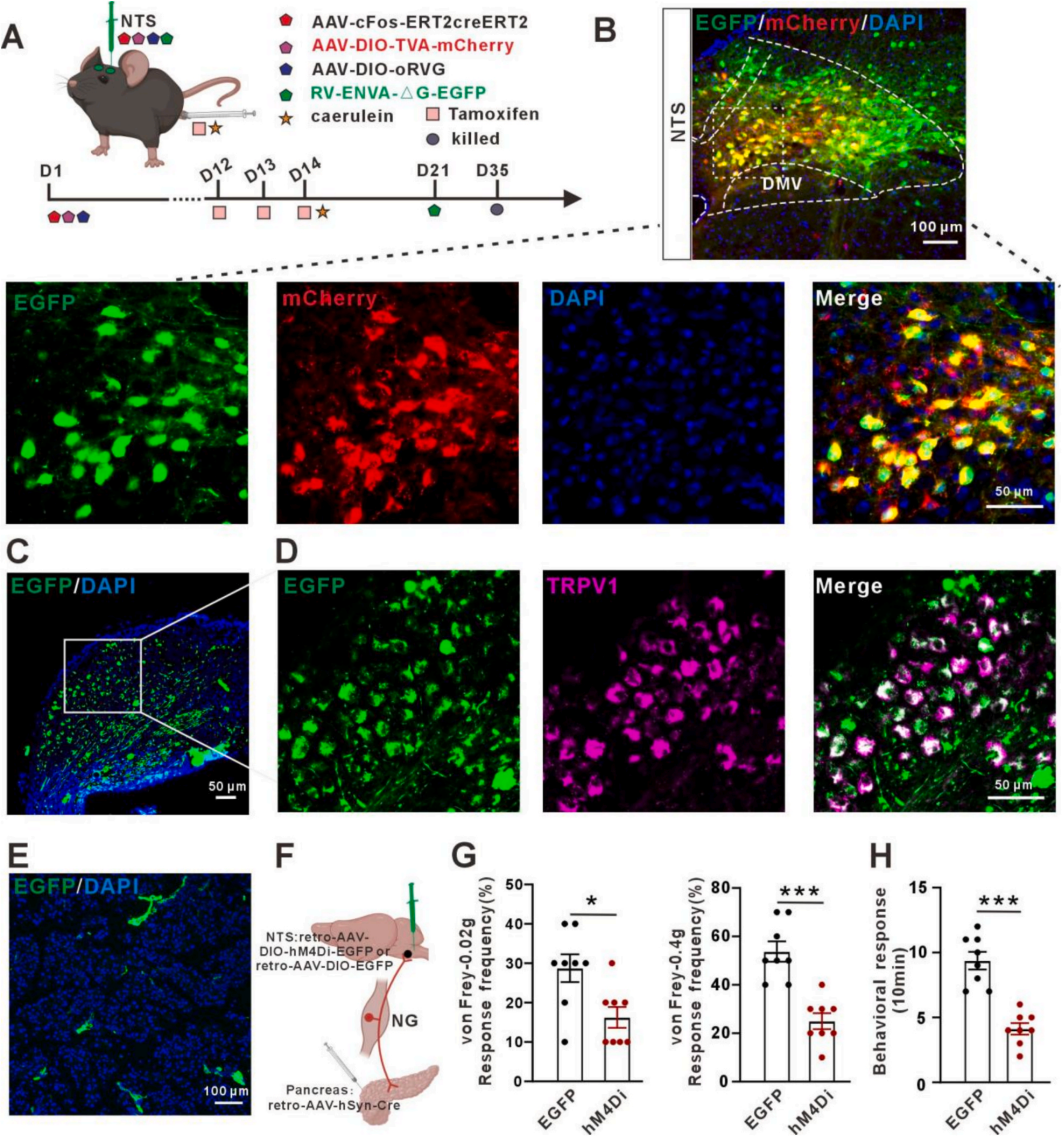
图4 迷走神经向孤束核传递内脏痛信号
为了探究急性胰腺炎时NTS神经元活性变化,采用Fos-creERT2策略,将AAV-cFos-ERT2creERT2与AAV-DIO-GCaMP6s共注射至NTS,腹腔注射他莫昔芬与雨蛙素诱导AP(图5A)。结果显示,NTS多个切面有大量GCaMP6s标记神经元(图5B),对照组仅见少量激活。cFos染色验证病毒标记效率:70.8%的GCaMP6s⁺细胞为cFos⁺,62.1%的cFos⁺细胞为GCaMP6s⁺(图5C-5E);光纤钙信号记录显示,雨蛙素处理后cFos⁺神经元活性显著升高(图5F)。采用Gi-DREADD抑制雨蛙素激活的cFos⁺神经元(图5G、5H),注射CNO后可显著缓解AP小鼠内脏痛(图5I-5J)。NTS含90%谷氨酸能(CaMKIIα⁺)和少量GABA能神经元,共染色显示GCaMP6s⁺神经元均与CaMKIIα共标(图5K-5L)。综上,NTS内谷氨酸能神经元激活介导AP内脏痛传入信号。
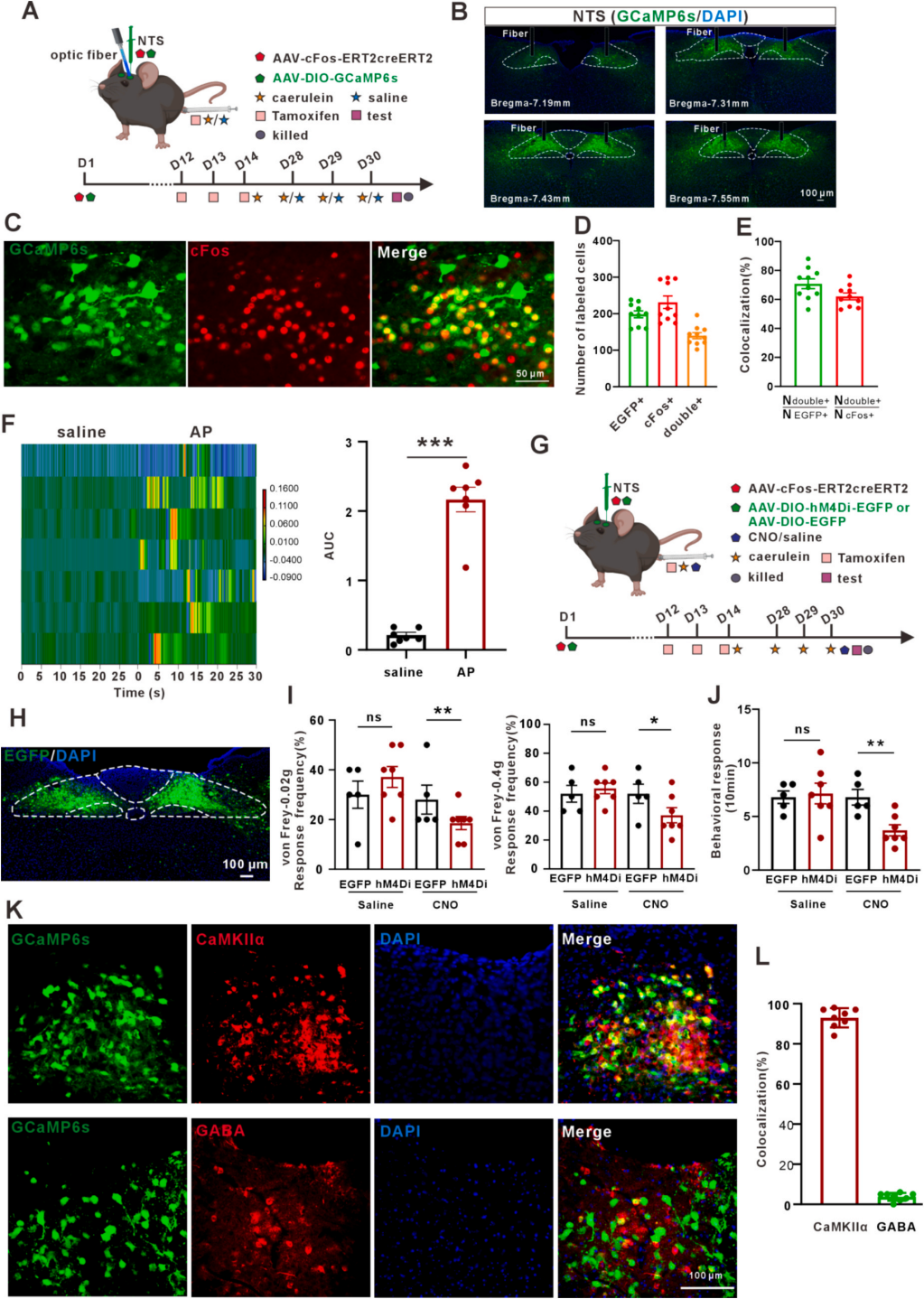
图5 孤束核NTSGlu调控急性胰腺炎内脏痛
探究NTSGlu神经元痛觉信号投射的高级脑区,将AAV-CaMKIIα-EGFP注入NTS绘制单突触投射图谱(图6A、6B)。示踪显示,其纤维末梢主要投射至下丘脑室旁核(PVN)、臂旁核(PB)、未定带(ZI)、丘脑室旁核(PVT)(图6C),均与痛觉感知相关。雨蛙素处理可显著上调PVN内cFos表达(图6D)。向NTS注射顺行病毒AAV2/1-hSyn-Cre、PVN注射Cre依赖型AAV-DIO-EGFP,21天后PVN可见NTS→PVN投射的绿色荧光。免疫荧光显示,EGFP信号与CaMKIIα大量共定位(图6E、6F);神经元亚型:3.6%为GABA阳性、10.1%为OXT阳性、5.8%为AVP阳性。接受NTSGlu投射的PVN谷氨酸能神经元在AP内脏痛中显著激活,提示PVN是NTS下游参与内脏痛加工的关键脑区。
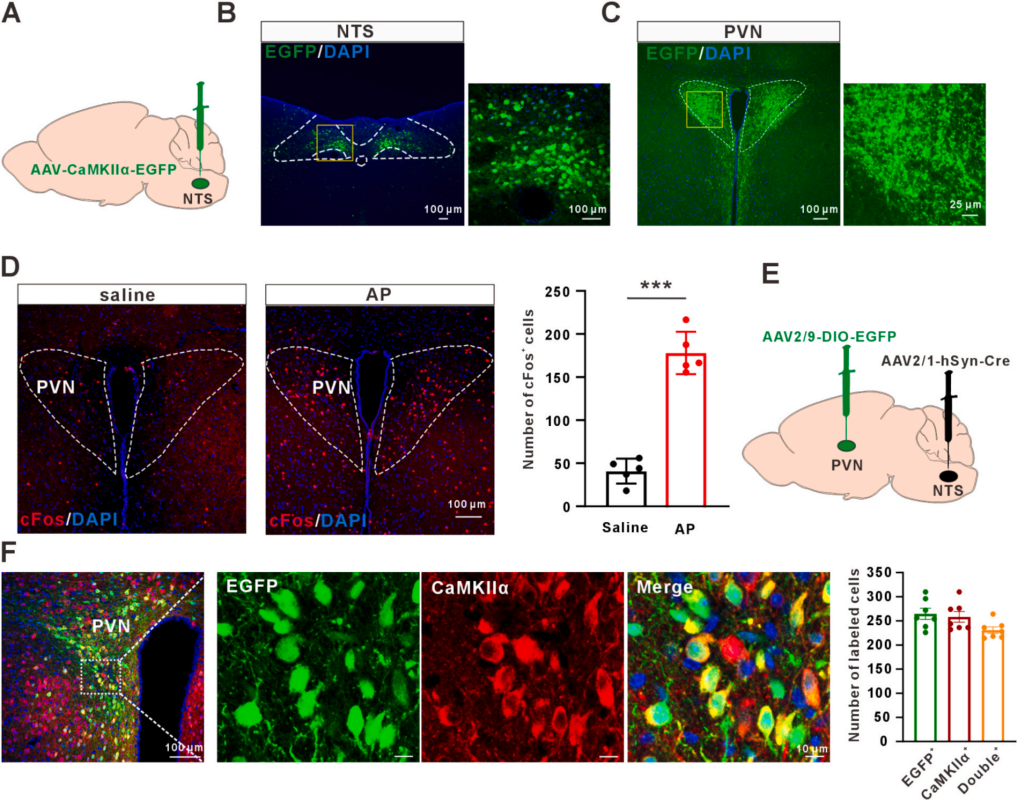
图6 投射至室旁核的孤束核谷氨酸能神经元
为阐明NTSGlu→PVNGlu通路在胰腺内脏痛中的作用,开展环路特异性调控实验。将AAV2/1-CaMKIIα-Cre与AAV-DIO-hM3Dq混合注射至NTS,AAV-DIO-GCaMP6s注射至PVN并植入光纤(图7A),腹腔注射CNO后记录钙信号,hM3Dq组PVNGlu神经元荧光强度显著高于对照组(图7B)。向NTS注射AAV2/1-CaMKIIα-Cre、PVN注射AAV-DIO-hM4Di-EGFP,腹腔给予CNO抑制该通路(图7C),可显著缓解AP诱导的内脏痛(图7D)。向NTSGlu末梢注射编码eNpHR的病毒进行光遗传抑制(图7E),也获得类似镇痛效果(图7F)。采用多重逆行示踪验证通路连接:胰腺注射retro-AAV-hSyn-mCherry,PVN注射retro-AAV-CaMKIIα-Cre,NTS注射Cre依赖性辅助病毒,三周后注射RV-ENVA-ΔG-EGFP(图7G)。NTS和NG内红绿荧光共定位,证实胰腺-大脑环路存在(图7H)。综上,胰腺-大脑轴(胰腺→NGTRPV1→NTSGlu→PVNGlu通路)介导AP后的内脏痛传入感知。
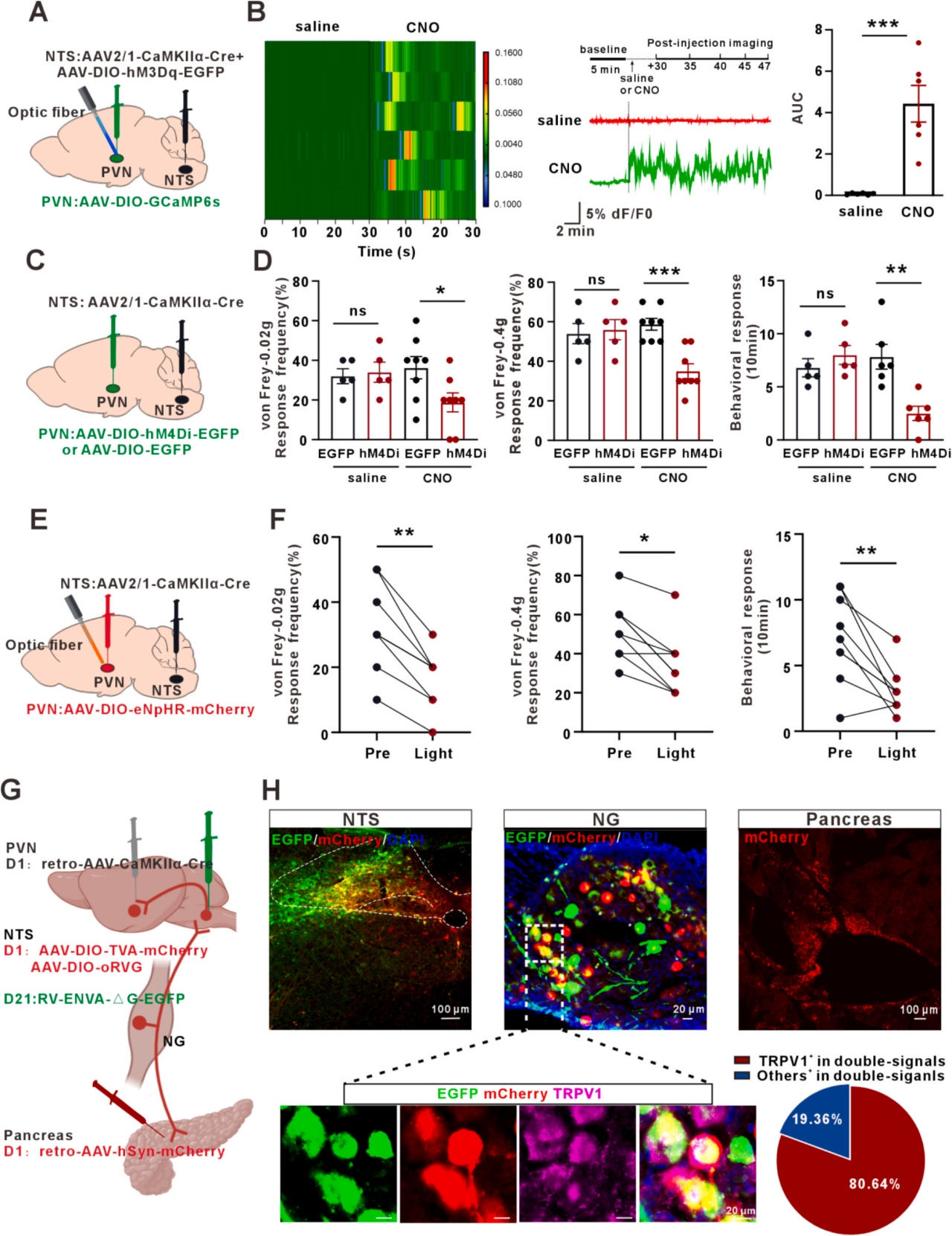
图7 NTSGlu→PVNGlu环路参与急性胰腺炎所致内脏痛
本文首次发现迷走神经介导的胰腺-大脑内脏痛通路(胰腺→NGTRPV1→NTSGlu→PVNGlu),该通路与经典的脊髓通路协同传递AP诱导的急性内脏痛信号。明确了该通路的关键分子与细胞节点:炎症因子TNF-α是外周敏化的核心因子,NGTRPV1、NTSGlu、PVNGlu是神经环路的关键节点,抑制任一节点均可有效缓解内脏痛。
本文使用的工具病毒布林凯斯均可提供:
同时布林凯斯也可提供各类定制服务请联系小布:18971216876(微信同号)或者咨询所在区域的销售经理获取更多信息。





