昼夜节律紧密调控摄食行为,慢性昼夜节律紊乱(如轮班工作、社交时差)会导致摄食时间紊乱,进而引发肥胖、2型糖尿病等代谢功能障碍。经典摄食与昼夜节律调控研究多聚焦于下丘脑区域(如下丘脑弓状核AgRP/NPY神经元),而腹侧海马体(vHPC)虽参与摄食调控,但其内NPY-INs在昼夜摄食中的作用及环路机制尚未明确。
2026年2月19日,上海交通大学徐天乐教授团队联合北卡罗来纳大学教堂山分校(UNC)宋娟教授团队,在Neuron期刊(IF=15)在线发表了题为“Ventral hippocampal NPY interneurons regulate circadian feeding in mice”的研究论文。在这项工作中,研究人员通过构建昼夜节律紊乱小鼠模型,结合化学遗传、钙成像、脑片电生理、脑区埋管给药和功能性磁共振成像(fMRI)等方法,揭示了腹侧海马表达神经肽Y的中间神经元(vHPC NPY-INs)通过下游脑区腹侧海马下托(vSub)的神经肽Y1受体(NPY1R)和神经肽Y2受体(NPY2R)实现对摄食昼夜节律的调控。这些发现不仅拓展了对摄食节律神经机制的理解,也为揭示昼夜节律紊乱相关的代谢性疾病的发病机制提供了新的理论依据。

作者首先构建了两种昼夜节律紊乱的小鼠模型,模拟现代生活中熬夜、倒班等导致的光暗周期异常,探究其对进食行为的影响。正常昼夜周期组、短期/长期LAN组(light at night夜间光照,24h亮灯)、长期DAD组(dark at day白天黑暗,24h黑暗),后续检测进食、代谢和体成分(图1A-B)。短期LAN对小鼠进食无显著影响,长期LAN会显著抑制小鼠暗期进食,导致整体进食节律紊乱(图1C-D)。长期DAD则表现为光期进食显著增加,同样破坏昼夜进食模式(图1E-F);长期LAN小鼠体重无显著变化,而长期DAD小鼠体重上升;但两者的脂肪、瘦肉含量均未发生明显改变,提示进食节律紊乱对体成分的影响存在模型特异性(图G-I)。
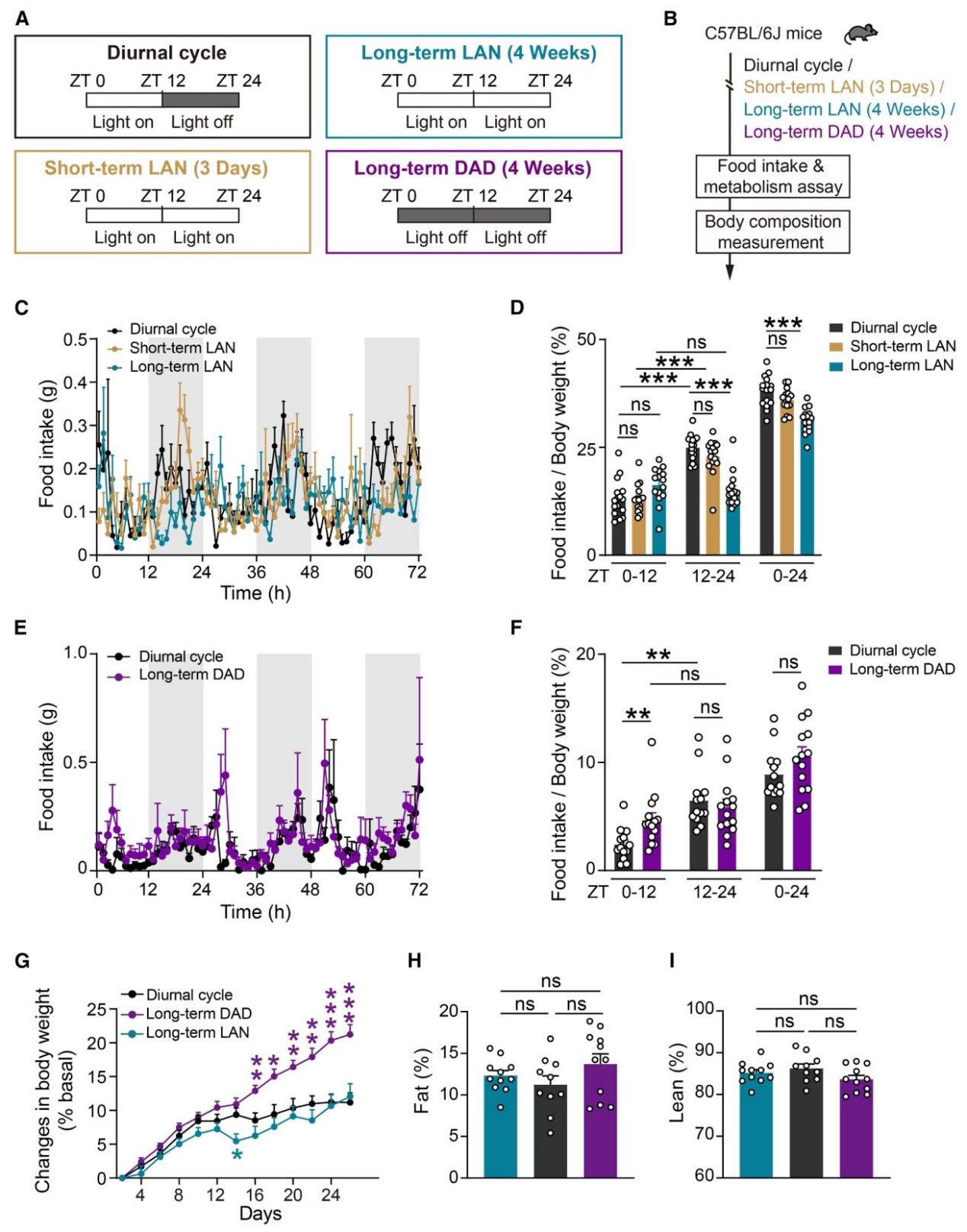
图1 | 长期LAN与DAD改变小鼠的摄食昼夜节律
在确认进食节律紊乱的表型后,作者通过活性标记和在体钙成像,筛选并验证了腹侧海马体NPY-INs的昼夜活动特征,这是研究的核心细胞靶点发现。利用c-Fos(神经元激活标记)与NPY双标染色,发现正常昼夜周期下,小鼠暗期(ZT14)vHPC NPY-INs的激活水平显著高于光期(ZT2);而长期LAN或DAD处理后,该昼夜激活差异完全消失(图2A-E)。通过光纤光度法在体记录钙信号,进一步验证:正常情况下vHPC NPY-INs的钙信号呈现暗期高、光期低的24h周期性波动;光期和暗期的钙事件频率存在显著差异,且该节律在昼夜紊乱模型中被破坏(图2F-J)。
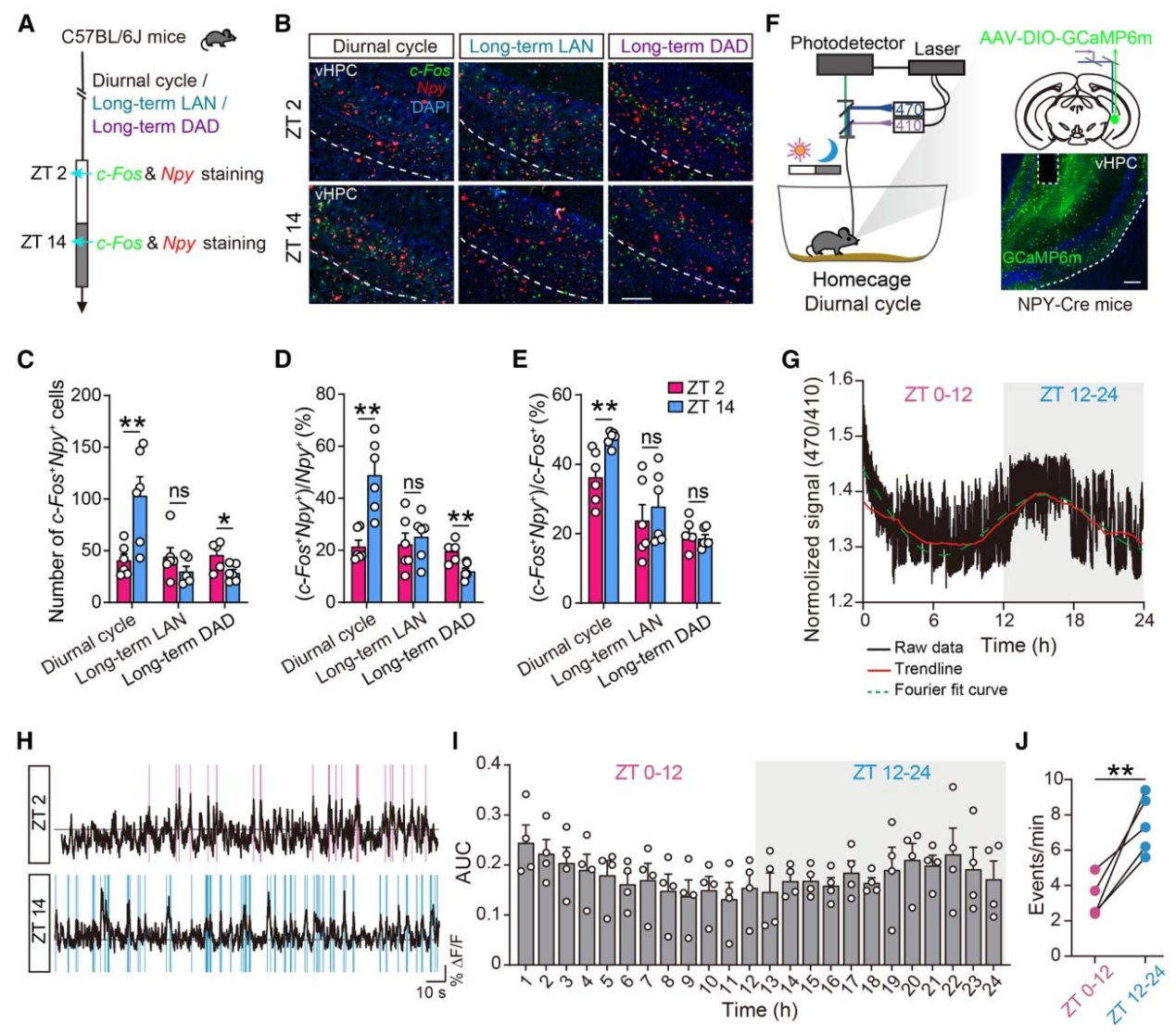
图2 | 昼夜节律紊乱破坏vHPC NPY-INs的节律性活动
为了证明vHPC NPY-INs的活性与进食行为之间的因果关系,作者利用Cre-loxP系统,在NPY-Cre小鼠的vHPC区域特异性表达兴奋性DREADD(hM3Dq)或抑制性DREADD(hM4Di),通过CNO给药调控神经元活性,检测进食变化。光期时,激活vHPC NPY-INs(hM3Dq)会显著增加进食量,抑制该神经元(hM4Di)则显著减少进食量;暗期时,该神经元本身处于高活性状态,激活无额外效应,而抑制则会显著减少暗期进食量;且该调控仅针对食物摄入,不影响饮水量(图3C-F)。长期激活vHPC NPY-INs不会改变小鼠的体重和体成分,原因是神经元激活同时增加了光期能量消耗,抵消了进食增加的效应(图3G-K)。此外,作者还通过多种行为学实验(新位置识别、新物体识别、Y迷宫、旷场实验等)排除了该神经元调控进食是通过影响记忆、焦虑等间接行为的可能性。
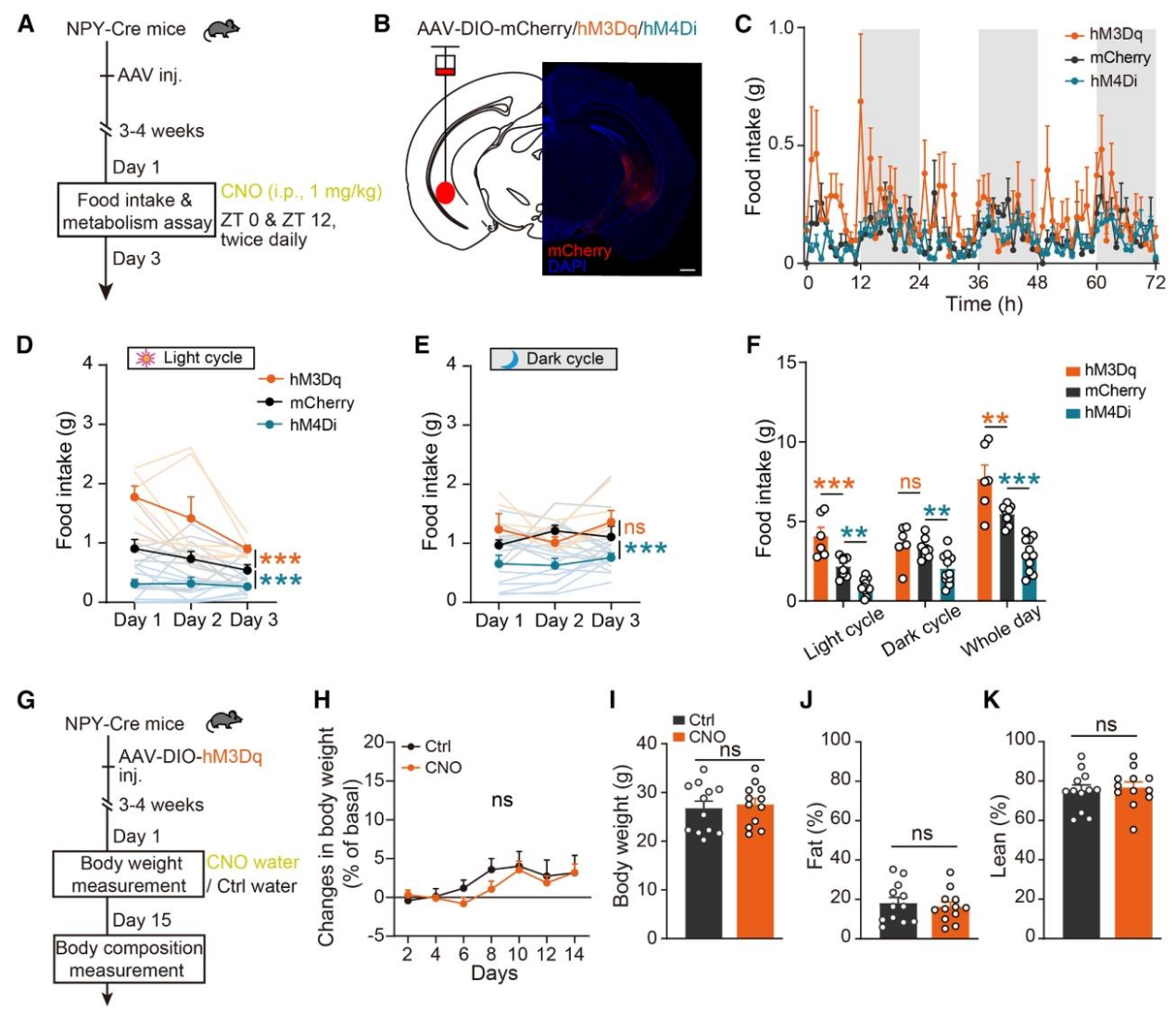
图3 | vHPC NPY-INs调控昼夜周期中的摄食行为
NPY-INs可共释放NPY和GABA两种信号分子,作者利用CRISPR-Cas9条件性敲除技术,分别敲除vHPC NPY-INs中的Npy基因和Slc32a1基因(GABA囊泡转运体,调控GABA释放),探究两种信号分子在昼夜进食中的特异性作用。Npy条件性敲除(cKO)后vHPC区域NPY的mRNA表达显著降低(图4A-D)。Npy cKO小鼠仅光期进食显著减少,暗期进食无任何变化(图4F-H)。Slc32a1 cKO效率验证,敲除后vHPC区域GABA转运体的mRNA表达显著降低(图4I-L)。Slc32a1 cKO小鼠仅暗期进食显著减少,光期进食无显著变化(图N-P)。vHPC NPY-INs存在昼夜信号切换机制——光期神经元低活性,主要通过NPY信号调控进食;暗期神经元高活性,主要通过GABA信号调控进食,两种信号分子在不同时段各司其职,共同维持昼夜进食节律。
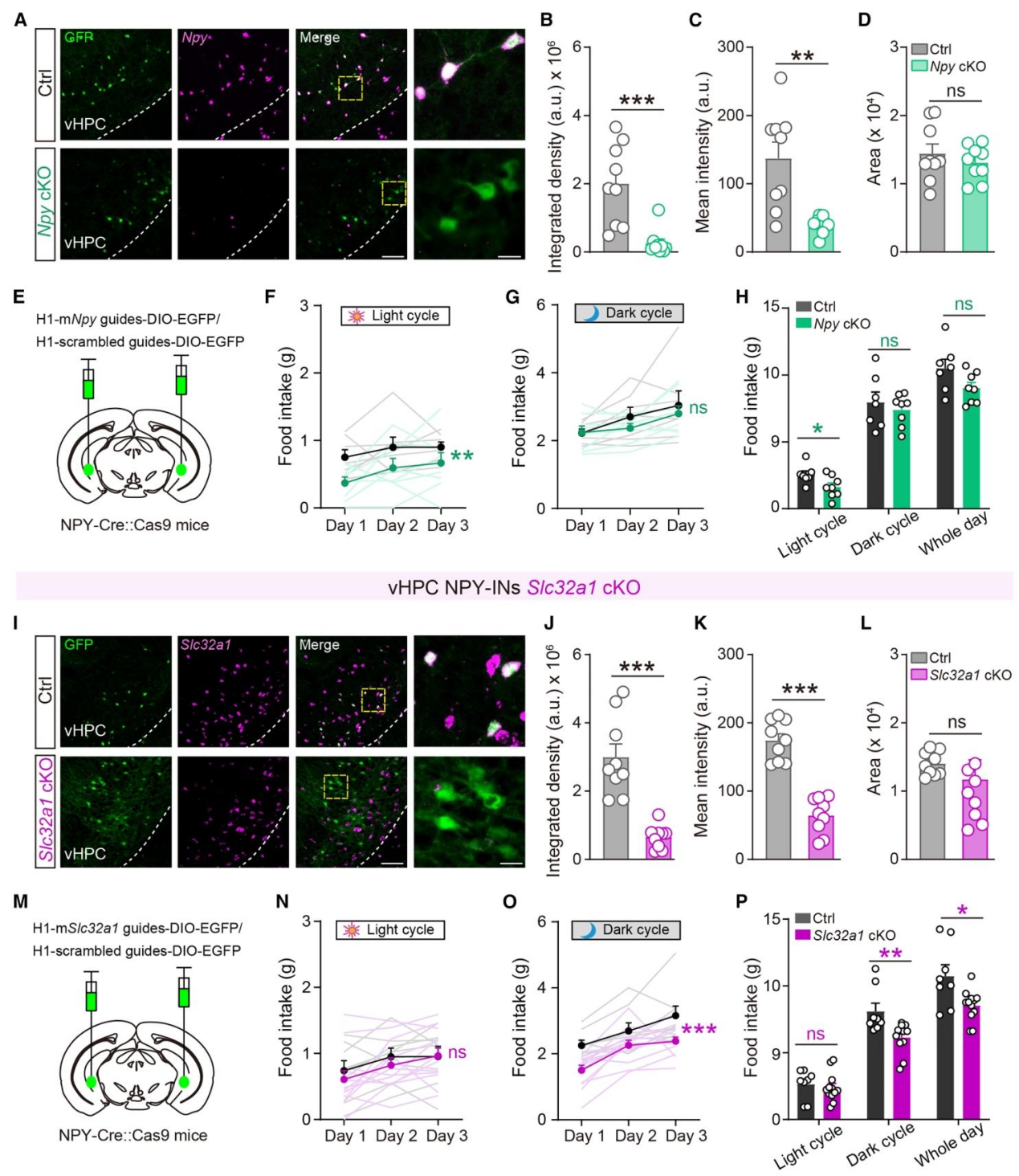
图4 | vHPC NPY-INs存在昼夜信号切换机制
明确了核心神经元和信号分子后,作者通过狂犬病毒逆向示踪筛选其上游输入核团,并结合电生理技术,解析了上游调控其昼夜活性的环路机制,这是研究的上游环路解析。狂犬病毒单突触逆向示踪发现,vHPC NPY-INs的主要上游输入核团包括内侧隔核(MS)、外侧内嗅皮层(LEnt)、背侧海马体(dHPC)和内侧视前区(MPOA),其中MPOA是经典的昼夜节律调控核团,成为研究重点(图5A-C)。RNAscope双标发现,正常昼夜周期下,MPOA的GABA能神经元(Vgat⁺)在光期激活水平更高,谷氨酸能神经元(Vglut2⁺)在暗期激活水平更高,与vHPC NPY-INs的光暗活性节律相呼应(图5D-K)。电生理记录发现,vHPC NPY-INs在光期的自发性兴奋性突触后电流(sEPSC)频率显著高于暗期,提示其突触前输入存在昼夜可塑性(图5L-O)。配对脉冲刺激实验显示,光期MPOA对vHPC NPY-INs的GABA能输入释放概率更高,暗期谷氨酸能输入释放概率更高;且96%以上的MPOA神经元可同时提供两种输入,通过调控兴奋/抑制(I/E)平衡,实现对vHPC NPY-INs昼夜活性的精准调控(图5P-W)。
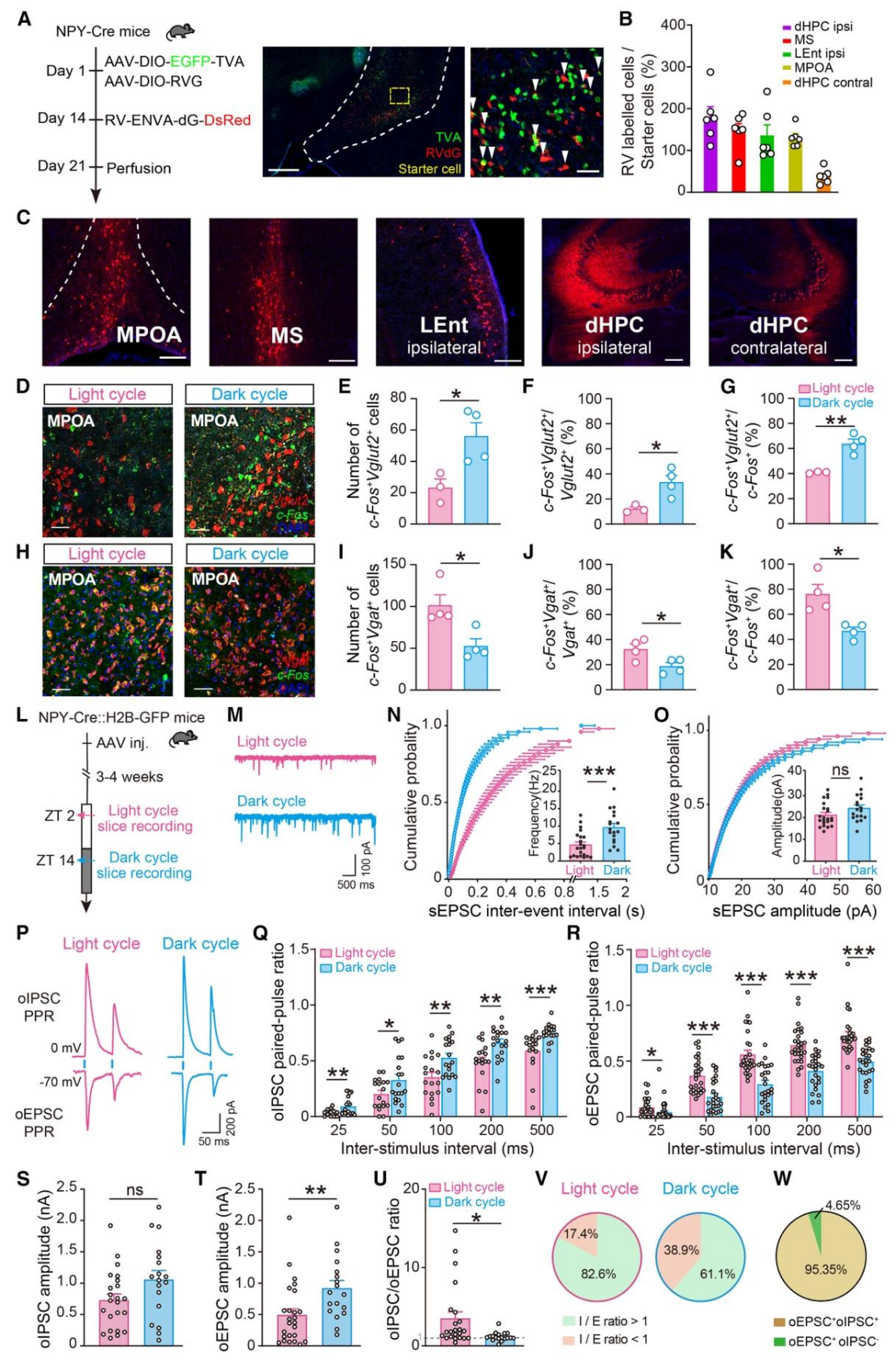
图5 | MPOA是调控vHPC NPY-INs昼夜活性的关键上游核团
为了验证MPOA→vHPC NPY-INs环路的功能因果关系,作者通过逆向病毒结合化学遗传学,特异性调控投射至vHPC的MPOA神经元,并进行环路交叉调控实验,验证上下游关系。特异性激活投射至vHPC的MPOA神经元,光期会增加小鼠进食(激活谷氨酸能输入,增强vHPC NPY-INs活性),暗期会减少进食(激活GABA能输入,抑制vHPC NPY-INs活性);抑制该投射则产生相反效应,而调控其他上游核团(MS/LEnt/dHPC)对进食无影响,证实MPOA的特异性(图A-D)。同时激活MPOA→vHPC投射并抑制vHPC NPY-INs,会完全阻断MPOA激活引起的光期进食增加,证实vHPC NPY-INs是MPOA调控进食的下游关键效应器(图E-H)。在NPY全身敲除小鼠中,激活MPOA→vHPC投射不再增加光期进食,反而使其减少,证实MPOA对光期进食的调控依赖vHPC NPY-INs释放的NPY信号(图I-L)。
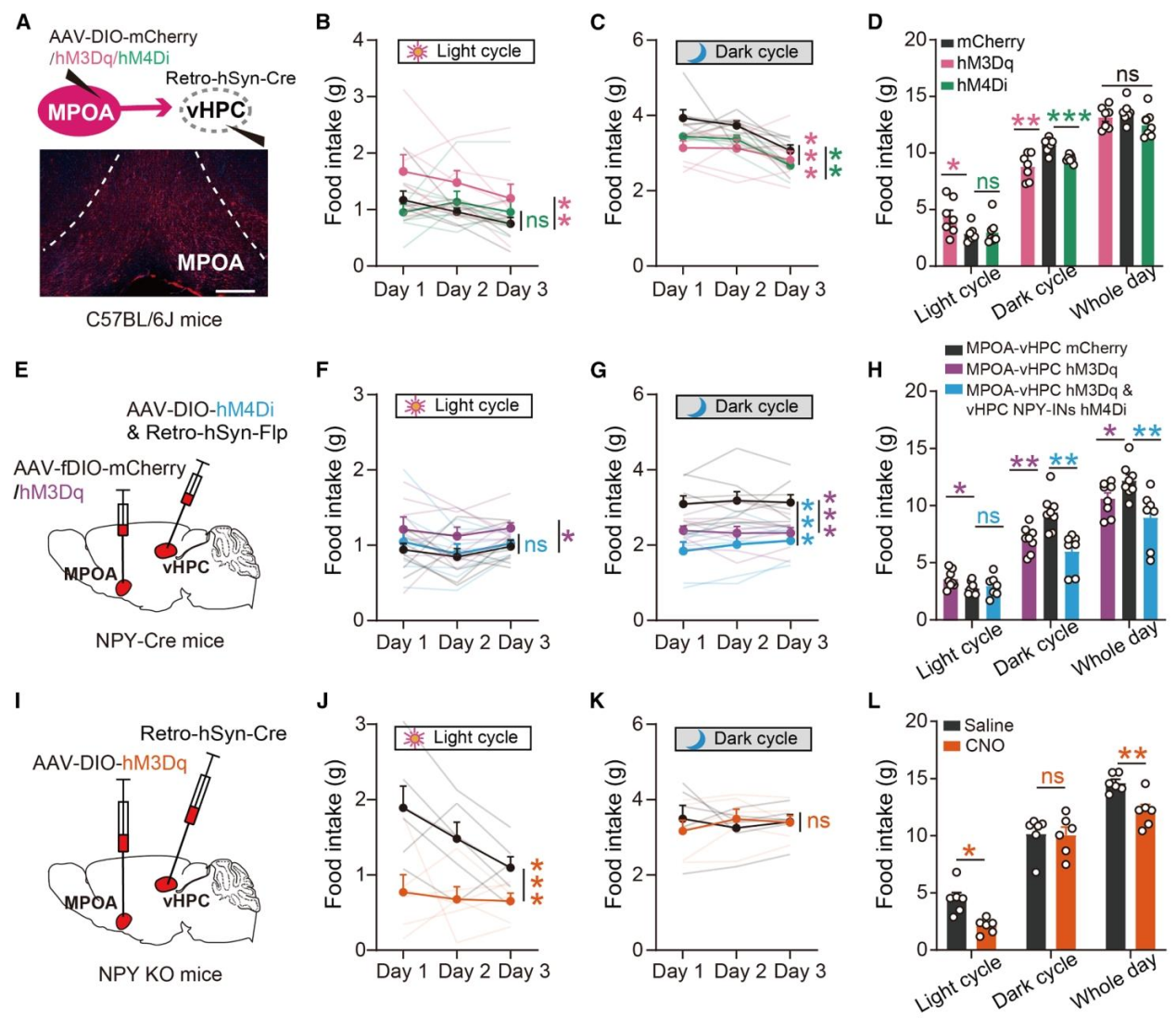
图6 | MPOA是vHPC NPY-INs调控小鼠昼夜进食的核心上游环路
解析完上游环路后,作者通过在体fMRI、顺行示踪、电生理和药理学干预,明确了vHPC NPY-INs调控进食的下游核团和受体机制,完成了整个环路的解析。清醒状态下fMRI发现,激活vHPC NPY-INs后,其与腹侧下托(vSub)的功能连接显著降低,提示vSub是其关键下游核团(图7A-E)。顺行示踪证实vHPC NPY-INs向vSub存在密集的神经投射,且vSub神经元中高表达NPY₁R和NPY₂R(NPY的主要受体)(图7F-G)。电生理记录发现,激活vHPC NPY-INs或直接施加NPY,会使vSub神经元发生超极化(抑制),而该效应可被NPY₁R+NPY₂R拮抗剂完全阻断,证实vHPC NPY-INs通过NPY抑制vSub神经元(图H-J)。侧脑室注射NPY₁R或NPY₂R拮抗剂,会完全阻断激活vHPC NPY-INs引起的光期进食增加,证实光期进食调控依赖vSub的NPY₁R/NPY₂R信号(图K-O)。同时激活vHPC NPY-INs和vSub神经元,会阻断vHPC NPY-INs激活引起的光期进食增加,证实vSub是vHPC NPY-INs调控光期进食的下游必要核团(图7P-S)。vHPC NPY-INs通过直接投射至vSub,释放NPY并结合其NPY₁R/NPY₂R受体,抑制vSub神经元活性,进而调控小鼠光期进食行为。
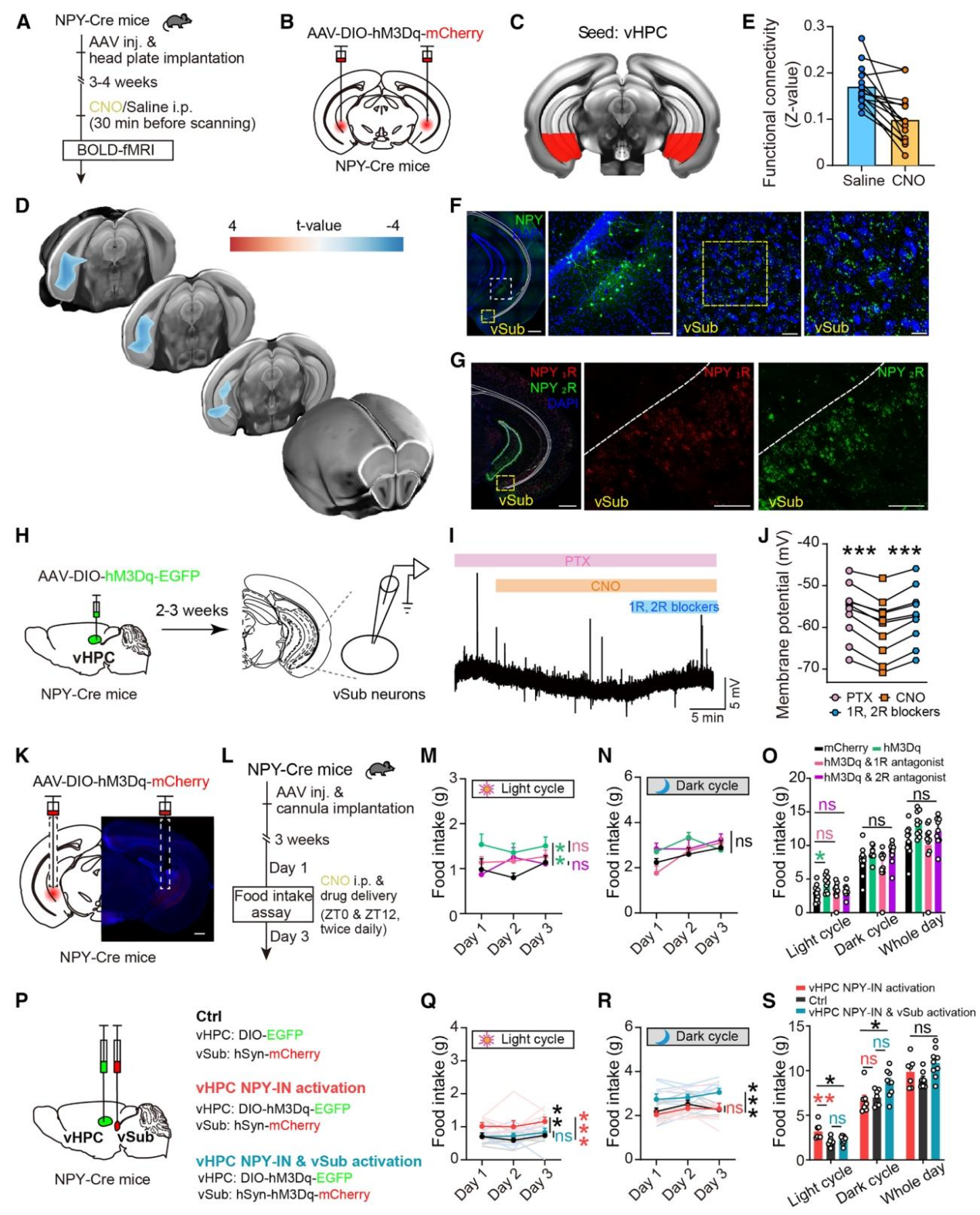
图7 | vHPC NPY-INs通过腹侧下托的NPY₁R和NPY₂R信号调控光周期摄食
结合所有实验结果,徐天乐教授团队首次描绘出调控小鼠昼夜进食的全新神经环路:MPOA(谷氨酸能/GABA能双重输入)→vHPC NPY-INs(光期NPY/暗期GABA信号切换)→vSub(NPY₁R/NPY₂R介导),该环路实现了从昼夜节律中枢到海马体,再到下游效应核团的精准调控,将经典的昼夜节律调控与海马体的高级神经功能结合,为进食节律调控提供了全新的视角。
本文使用的工具病毒布林凯斯均可提供:
同时布林凯斯也可提供各类定制服务请联系小布:18971216876(微信同号)或者咨询所在区域的销售经理获取更多信息。





